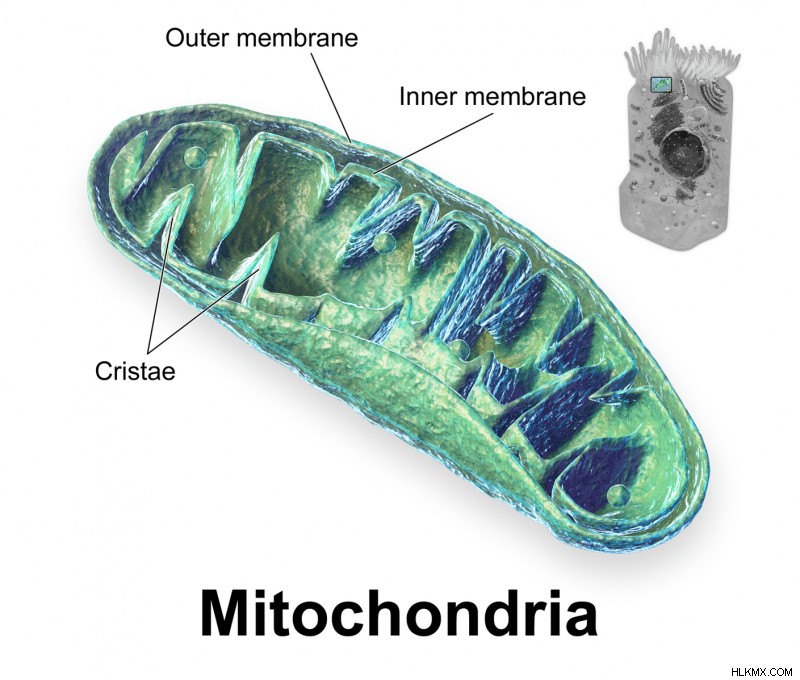
미토콘드리아는 세포의 강국 소기관입니다. 이들은 주로 다양한 세포 활성에 사용되는 고 에너지 분자 인 ATP를 생성 할 책임이있다. ATP 생산은 호흡 과정에서 발생합니다. 작은 구획 내에서 발생했습니다.
그러나, 미토콘드리아를 만들고 활성에 필요한 대부분의 단백질은 내부에서 합성되지 않고 오히려 세포 단백질의 대부분과 마찬가지로 세포질에 위치한 리보솜이라고 불리는 단백질-합성 기계에 의해 합성된다. 문제는 미토콘드리아를 구축하는 데 필요한 단백질이 어떻게 작기에 들어가는 일을 하는가? 그 대답은 미토콘드리아가 개발 한 특수 단백질 수입 메커니즘에 있습니다.
미토콘드리아는 수십억 년 전에 조상 진핵 세포에 의해 산소 소비 원핵 생물의 혼입으로 인한 것으로 생각된다. 내 생물 생물학적 과정이라고 불리는이 진화는 곰팡이, 식물 및 동물과 같은 현대 진핵 생물을 일으킨 주요 전환점입니다. 그러나 식물은 하나의 내 endosymbiotally 유래 소기관, 미토콘드리아 및 플라 스티드를 가지고 있기 때문에 다른 사람들에 비해 독특한 특징을 가지고 있습니다. 둘 다 주로 생활에 필요한 에너지와 분자를 생성하는 책임이 있습니다. 흥미롭게도, 세포질에서 합성 된 일부 단백질은 미토콘드리아 및 플라 스티드 둘 다를 대상으로 할 수있다. 결과적으로, 식물의 미토콘드리아 성분 중 일부는 독점적이며 식물 분자 생물 학자들 사이에서 명명을 밝히는 데 상당한 관심을 갖게되었습니다.
작은 크기에도 불구하고, 미토콘드리아의 단백질-표적화 메커니즘은 놀랍게도 복잡하며 다양한 단백질 복합체를 포함한다. 항상 매우 바쁘다! 미토콘드리아는 이중 막 결합 소기관이다. 결과적으로, 단백질 트랜스 코스의 두 가지 주요 성분이 외부 막에서 내부 막에서 내부 막에서 내부 막 (IMS)까지의 단백질을 이동시켜 단백질을 외부 및 내부 막에 존재한다.
외막에는 외부 막 (TOM) 및 정렬 및 조립 기계 (SAM) 복합체가 초기에 표적화 된 단백질 및 기타 거대 분자를 인식하는 데 도움이된다. 내부 막에서, 내부 막 (TIM) 복합체의 전산물은 단백질 선택기로서 작용한다. IMS에서, 미토콘드리아 내부 막 공간 수입 및 어셈블리 (MIA) 복합체에 의해 일반적으로 수행되는 단백질 분류. 식물 미토콘드리아 수입 성분은 그림에서 볼 수 있습니다. 함께, 이들 국토 케이스 성분은 서로 다른 특징을 가진 표적화 된 단백질을 전위하는 데있어 4 가지 주요 경로를 구성한다. 이것들은 일반적인 수입 경로, 캐리어 수입 경로, MIA 경로 및 SAM 경로입니다. 주로 사전 시퀀스, 내부 신호를 갖는 단백질, 트윈-시스테인 단백질 및 β- 배럴 단백질을 각각 수송하는 데 책임이있는 SAM 경로입니다.
.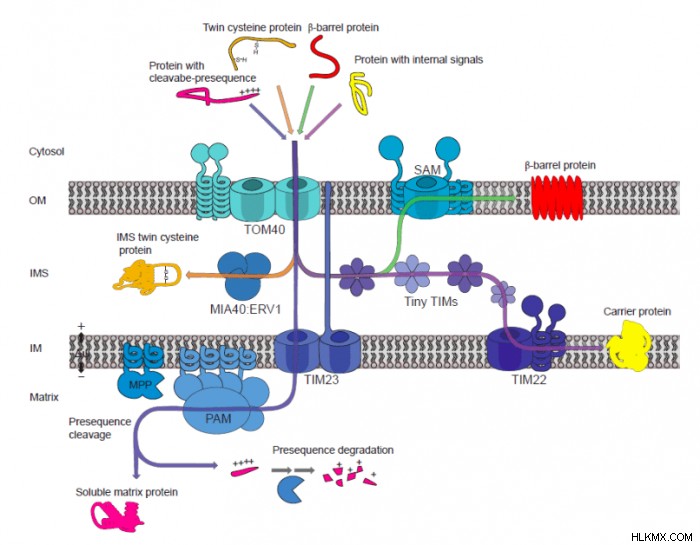
이들 복합체는 신호를 식별하고 단백질을 다른 소기관 위치로 운반하는 데 함께 작용하는 다수의 단백질 부분을 가지고있다. 흥미롭게도, 모든 진핵 생물에서 국군 제 복합체가 관찰 되었음에도 불구하고, 식물에서 수입 성분의 성분 및 구조적 배열은 다른 것과 유의하게 다를 수있다. 예를 들어, 일부 Tom 성분은 식물에서 발견되지 않았지만 효모에서 특성화되었습니다 ( Saccharomyces cerevisiae ) Tom70 및 Tom22/9와 같은)
반면에, 일부 성분은 무엇보다도 ATP 신타 제의 서브 유닛의 수송을 담당하는 OM64와 같은 식물에만 국한된다. ATP 신타 제는 미토콘드리아의 주요 역할 인 ATP를 생산하는 효소 복합체이다. OM64의 다른 필수 톰 성분과 함께 OM64의 결실은 식물에 치명적이며 미토콘드리아 생물 생성 및 활동에서의 필수 역할을 보여줍니다.
애기 장대 미토콘드리아 수입 성분에 대한 최근의 체계적인 단백질 학 연구에 따르면 모든 구성 요소가 동일한 회전율을 갖는 것은 아닙니다. 일부는 이직률이 상대적으로 빠릅니다. 이는 풍부도 변화가 다른 중요한 구성 요소보다 높음을 의미합니다. 이러한 "빠른"가져 오기 구성 요소에는 TOM20 및 TIM17이 포함됩니다. 이러한 "빠른"성분은 빠른 단백질 합성을 필요로하는 식물의 성장 및 발달 단계에 필요하다는 것이 예측된다. TIM17 상 동체는 실제로 식물 발아 및 스트레스 반응에 중추적이라는 것을 언급 할 가치가 있습니다.
단백질 수입의 이러한 복잡한 배열은 다음 질문을 제기합니다. 이러한 복합체는 어떤 단백질이 어떤 단백질에 들어갈 수 있고 어떤 단백질이 특정 위치로 가야하는지 인식하는 방법을 어떻게 인식합니까? 이 수입의 핵심은 단백질 서열 내의 표적화 신호에있다.
표적화 펩티드는 단백질 수입 성분에 의한 인식 및 분류에 해당하는 표적화 된 단백질 내의 아미노산 서열 세트이다. 타겟팅 신호에는 주로 두 가지 유형이 있습니다 :절단 가능하고 절약 할 수 없습니다. 미토콘드리아-정열 된 단백질의 대다수 (약 70%)는 수입시 절단되고 다단계 처리 경로를 통해 아미노산으로 추가로 소화 될 예정인 펩티드 (presequence)라고 불리는 절단 가능한 표적화 펩티드를 함유한다. 최근의 연구는 표적화 펩티드의 2 차 구조가 서열이 아닌 수입 성분에 의한 인식을 결정한다는 것을 보여 주었다. 예를 들어, 미토콘드리아 presequences는 종종 Tom Complex에 의해 인식되는 양친 매성 α- 헬리스를 형성합니다. 이것은 왜 일부 단백질이 정확한 시퀀스보다는 구조적 특징을 인식하기 위해 트랜스 코카이스의 선호로 인한 다중 소기관을 표적으로하는지 설명 할 수 있습니다.
.식물 미토콘드리아 단백질 수입 성분은 다른 진핵 생물에 비해 낮은 보존을 갖는다. 뚜렷한 단백질 수송 성분은 다른 진핵 생물의 밀접한 관련 단백질과 구조적으로 그리고 조절하는 현재까지 특성화되었다. 따라서, 서열 분석 및 계통 발생학 연구에서만 새로운 수입 구성 요소를 특성화하는 것은 어렵다. 유전학 및 생화학 적 연구는 미토콘드리아 생물 생성 및 활동의 게이트 키퍼, 식물의 성장 및 스트레스 반응 규정으로 작용하는 이러한 새로운 수입 구성 요소를 특성화하는 데 빛을 비출 수 있습니다.
이러한 결과는 Plant Mitochondrial Protein 수입 구성 요소 :The INS and Outs라는 제목의 기사에 설명되어 있으며, 최근 Biochemical Journal 에 발표되었습니다. . 이 작업은 Mabel LA의 Abi S. Ghifari에 의해 수행되었습니다. 웨스턴 호주 대학교의 Gill-Hille 및 Monika W. Murcha.