과거는 중요하고 놀라운 유산을 남길 수 있습니다
자연적으로 또는 본질적으로 수정되지 않은 상태에서 귀중한 식생 유형의 넓은 연결된 영역을 보존하는 것은 기후가 빠르게 변화하는시기에 계획 당국에 복잡한 도전입니다. 고생물 생태 학적 데이터는 산림 연속성, 종 다양성, 유형 및 교란의 빈도, 인위적인 영향의 정도에 영향을 미쳤던 이전 세대의 나무에서 과거의 유산에 빛을 비출 수 있으며, 현재 지역 사회를 위조하고 미래 계획을 세운 상황을 더 잘 이해하게됩니다. 그러나 지배적 인 나무의 기대 수명이 일반적으로 훨씬 더 길어도 시간 규모에 걸친 생태 과정을 기록하는 데이터 세트는 거의 사용되지 않습니다 (Willis and Bhagwat, 2010).
과거의 교란의 유산을 이해하는 것은 유럽의 온대와 대서 림에서 특히 중요하며, 농업을위한 산림 제거와 자연 동물 탐색 및 화재 체제의 중단이 적어도 신석기 시대부터 널리 기록되었습니다 (Whitlock et al. 2017). 일부 열대 산림 사례 연구에도 불구하고 전 세계의 다른 지역의 산림에 상당한 인위적 영향의시기는 비슷하거나 젊습니다. 비록 일부 열대 산림 사례 연구에도 불구하고 선사 시대의 인간 활동이 한때 믿었던 것보다 훨씬 광범위하다고 제안했지만 (Willis et al. 2018).
.지속적인 Holocene Forest Cover> 13,500 년이있는 스웨덴 사이트는 "자연"및 인위적 교란 체제에 의해 주도되는 시간을 통해 상당한 변화를 보여줍니다
현재 "핫스팟"자연 보호 구역 인 Almeberget의 주요 생물 다양성 값은 스웨덴 남서부 (커버 사진)의 희귀 이끼, 곰팡이, 브라이오 피 테스 및 너도밤 나무 ( fagus sylvatica )를 포함합니다. ),이“키스톤 트리”의 지배력은 지난 마지막 C. 13,500 년 (그림 1). 꽃가루와 숯 데이터를 사용하여 재구성 된 식생 및 화재 이력, 변화하는 종의 조합과 자연 및 인위적 교란의 복잡한 혼합물을 보여줍니다.
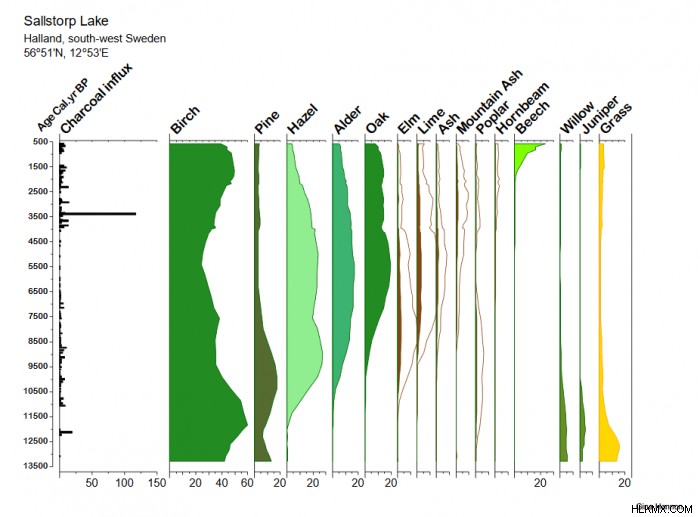
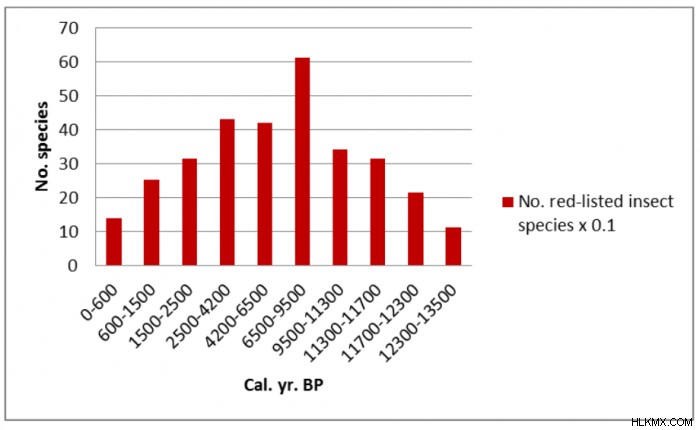
장기 트렌드는 시간이 지남에 따라 다양한 변동성이 있으며, 현재 관리를위한 목표로 사용할 단일 기준선 또는 기준점이없는 다양한 변동성이 있음을 보여줍니다 (Bradshaw et al. 2015; Hannon et al. 2018; Hannon, 2019). 9500에서 6500 Cal 사이의 초기 혼합 숲. 저 강도 화재가 자주 발생하는 YR BP는이 부위에서 가장 다양한 레드리스트 곤충 종과 관련이 있습니다 (그림 2). 비교적 저 강도 인 인위적 교란과 함께 숲 연속성과 구형 낙엽수의 조각난 저수지 개체군은 현재, 종이 풍부한, 희귀 한 epiphytic flora, Wood-inhabiting 곰팡이 및 비간 뇌물 공동체의 생존에 중요했을 가능성이 높습니다 (Hannon et al, 2018). 이러한 많은 산림 조각들은 미래의 인위적 영향과 기후 변화에 더 취약해질 수 있습니다.
최근 스칸디나비아 남부의 산림의 생물 균질화
오히려 순수한 너도밤 나무 숲은 트리 커버가 긴 연속성을 가진 부지에 서 있습니다. 최근의 광범위한 생물 균질화는 상업용 임업 운영을 포함한 자연 및 문화적 세력에 의해 주도되었다 (Seppä et al. 2009; Lindbladh et al. 2014). 스웨덴 남부의 다양한 낙엽 숲을 가문비 나무 단일 분류 ( picea abies )는 오크에 의존하는 많은 곤충, 브라이오 피트 및 이끼의 국소 손실과 관련이 있습니다 ( quercus ), 라임 ( 틸리아 ), 포플러 ( populus ), Alder ( alnus ) 및 기타 나무 (Jonsell et al. 1998). 북유럽을 통한 너도밤 나무의 후기 홀로 세인 스프레드는 독일과 덴마크의 넓은 숲 전체에서 나무 다양성이 감소했으며, 산림 구성의 이러한 변화는 자연적이고 인위적인 요인의 조합에 의해 주도되었습니다 (Giesecke et al. 2011; Bradshaw et al. 2010; Bradshaw et al. 2015). 가문비 나무와 소나무가있는 대규모 심기 프로그램 ( pinus ) 1900 년대 초부터 화재 감소와 함께 균질화 과정을 가속화하여 자연 보존을위한 위기를 일으켰습니다.
스웨덴의 관리 우선 순위는 현재 종 구성의 영향을 미칩니다
스웨덴의 산림 매장량과 국립 공원의 보존은 종종 위협받는“빨간색”종의 존재에 의해 안내됩니다 (Brunet et al. 2010; Fritz, 2011). 산림의 보호는 일반적으로 핵심 지역 내에서의 개입이없고 주변 완충 구역에서 죽은 목재와 같은 다양한 적합한 기질의 격려를 포함하여 적색 목록 및 기타 산림 종의 확산을 용이하게합니다 (Halme et al. 2013). 이 접근법은 산림 생물 다양성의 중요한 구성 요소의 장기 유지를위한 교란 역학의 중요성을 과소 평가할 수 있습니다. 현재 스웨덴 남서부의 보호 된 숲은 과거보다 나무 다양성이 낮으며, 적수 종의 더 작은 개체수가있을 가능성이 높습니다 (그림 2). 이 스탠드는 보존에 대한 자연스러운 초점이지만 과거에 대한 지식은 이전의 더 풍부한 시스템의 살아남은 잔재를 보호하는 목표를 넘어서서 활발한 복원 조치를 제안하는 데 사용될 수 있습니다.
.장기 데이터의 통찰력은 산림 생물 다양성 보존을위한 새로운 조치를 제안합니다
고생물학적 정보는 보존 관리를위한 새로운 권고의 기초를 형성 할 수있다. 현대 공동체와 비교됩니다. 유리한 조건을 촉진하고 미래를위한 실용적인 목표를 설정하기위한 관리 체제에 통합됩니다 (Jackson and Hobbs, 2009; Anderson et al. 2006; Dietl et al. 2015). 특히, 현재의 식생 공동체가 초기 조건에서 어떻게 발전했는지에 대한 지식과 미래의 구성이 발전 할 수있는 것들에 대한 지식은 실용적이고 성공적인 산림 보존 조치를 계획하는 데 관리자에게 가치가 있습니다. 스웨덴의 Halland 카운티는 해당 지역의 5.6%가 자연 보호 구역에서 보호되었으며 Palaeocologists에게 적절한 관리 계획에 기여하도록 의뢰했습니다.
우리의 분석 결과 (그림 1)는 고생물학적 데이터가 과거의 변화를 자세히 설명하는 설명 적 기록 일뿐 만 아니라, 오늘날의 생태계 관리와 관련된 종 구성 및 과정에 대한 정보를 제공한다는 것을 보여 주었다. 이는 여러 관리 조치의 구현에 기여할 수 있습니다. 너도밤 나무 나 가문비 나무의 단일 문화 근처에 의해 지배되는 넓은 산림 지역은 더 긴 시간 관점에서 매우 이례적이며 잠재적으로 급속한 기후 변화의시기에 탄력성이 부족합니다.
이전의 혼합 낙엽 숲의 복원은 생물 다양성을 증가시킬 것이며 라임, 오크, 엘름 및 알더와 같은 나무는 현재 기후에서 지속 가능한 인구를 건설 할 수 있어야합니다. 그들의 성공은 브라우징 체제의 관리와 사슴 개체군의 통제에 달려있다. 현재의 기후 경향으로 화재 위험이 증가 할 가능성이 있지만 과거의 혼합 낙엽 산림은 가문비 나무와 너도밤 나무에 불리하게 될 온화한 지상 화재 체제를 용납했습니다. 연구 의제는 보호 된 준비금 개념을 넘어서서 생물 다양성에 대한 주요 위협이 될 수있는 미래 기후에 적합한 새로운 생태계를 고려해야합니다 (Willis and Bhagwat, 2010).
.참조 :
- Anderson, N.J., Bugmann, H., Dearing, J.A. 및 Gaillard, M-J. 2006. Palaeoenvironmental 데이터 및 모델 연결 과거를 이해하고 미래를 예측하기 위해 연결. 생태 및 진화 동향 21 (12), 696–704.
- Bradshaw, R.H.W., Lindbladh, M. 및 Hannon, G.E. 2010. Holocene 후반 스칸디나비아 숲에서 화재의 역할. International Journal of Wildland Fire 19, 1040–1049.
- Bradshaw, R.H.W., Jones, C.S., Edwards, S. J. 및 Hannon, G. E. 2015. 서유럽의 산림 연속성 및 보존 가치. Holocene 25 (1), 194–202.
- Brunet, J., Fritz, Ö 및 Richnau, G. 2010. 유럽 너도밤 나무 산림의 생물 다양성 - 지속 가능한 산림 관리를위한 권장 사항이있는 검토. 생태 게시판 53, 77–94.
- Dietl, G.P., Kidwell, S.M., Brenner, M., Burney, D.A. et al. 2015. 보존 고생물학 :과거에 대한 지식을 활용하여 보존 및 복원에 정보를 제공합니다. 지구 및 행성 과학의 연례 검토 43, 79–103.
- fritz, Ö. 2011. Lunglav Minskar och Bokfjädermossa Ökar I Hallands Bokskogar. Svensk Botaniskt Tidskrift 105, 163–177.
- Giesecke, T., Bennett, K.D., Birks, H.J.B., Bjune, A.E. et al. 2011. Holocene 식물의 속도는 동기 개발에 대한 테스트를 변화시킵니다. 4 차 과학 검토 30 (19–20), 2805–2814.
- Halme, P., Allen, K.A., Auniņš, A., Bradshaw, R.H.W. et al. 2013. 생태 복원의 도전 :북유럽의 숲에서의 교훈. 생물학적 보존 167, 248–256.
- Hannon, G.E. 2019. Almeberget Nature Reserve의 장기 프로세스 :A 'Hotspot'높은 보존 지분의 숲. Halland County 당국에 대한 보고서. (프레스에서).
- Hannon, G.E., Halsall, K.M., Molinari, C. et al. 2018. SW 스웨덴에서 지난 13,500 년 동안 과거 산림 역학의 재건. Holocene 28 (11), 1791–1800.
- 잭슨, S.T. &Hobbs, R.J. 2009. 생태 역사에 비추어 생태 복원. 과학 325 (5940), 567–569.
- Jonsell M., Weslien J. 및 Ehnström B. 1998. 스웨덴의 적색 실린 사파 실산 무척추 동물의 기질 요구 사항. 생물 다양성 및 보존 7 (6), 749–764.
- Lindbladh, M., Axelsson, A-L., Hultberg, T. et al. 2014. Broadleaves
에서 가문비 나무로 - 스웨덴 남부의 대담한. 스칸디나비아 산림 연구 저널 29 (7), 686–696. - Seppä, H., Alenius, T., Muukkonen, P. et al. 2009. 정량적 나무 바이오 매스 재구성의 기초로서 교정 꽃가루 축적 률. Holocene 19, 209–220.
- Whitlock, C., Colombaroli, D., Conedera, M. &Tinner, W. (2017). 산림 보존 및 관리를위한 안내서로서의 토지 이용 역사. 보존 생물학 32 (1), 84–97.
- 윌리스 K.J. 및 Bhagwat, S.A. 2010. 생물학적 다양성 보존에 중요한 문제 :과거의 답변. 과거의 환경 6 (6), 759-769.
- Willis, K.J., Jeffers, E.S and Tovar, C. 2018. 지상 생태계를 탄생하는 이유는 무엇입니까? 과학 359 (6379), 988-989.