석유는 전 세계의 주요 에너지 원을 구성합니다. 세계 인구의 지속적인 증가로 인한이 자원의 과도한 소비로 인해 석유 매장량이 고갈되었으며, 이는 온실 가스 배출이 방출되어 기후 변화의 영향이 증가함에 따라 (Stephanopoulos, 2007).
이러한 문제에 대한 잠재적 인 해결책은 바이오 연료, 옥수수, 포플러 및 스위치 그라스와 같은 식물에서 생성 될 수있는 지속 가능한 에너지의 대체 공급원에 있습니다.
엔진에 사용 된 최초의 주요 대안 바이오 연료는 1990 년대 초에 에탄올로 일반적으로 알려진 바이오 에탄올이었으며, 이는 휘발유와 혼합되어 10% 에탄올과 90% 가솔린 E10 연료를 생성 하였다. 다른 혼합물에는 최대 15% 에탄올 및 E85를 포함하는 E15 및 유연한 연료 차량에 사용할 수있는 E15가 포함됩니다. 오늘날 미국에서 사용되는 연료의 98% 이상이 E10 혼합 연료이며, 이는 일반적이고 편리한 연료 대안입니다.
그러나 이것은 가장 효율적인 재생 가능한 연료가 아닙니다. 에탄올은 바이오 연료 사용 우선 순위에 대한 탄탄한 출발점을 제공했지만,이 특정 바이오 연료는 고유의 과제를 가지고있어 저장된 에너지와 부식 효과를 포함하여 엔진 부품을 제조하는 데 사용되는 철강의 균열을 초래하는 연비로의 추가 확장을 제한합니다 (Clemente, 2015).
결과적으로, 연구자들은 일반적인 박테리아에서 유전자 공학을 사용하여보다 효율적인 대체 바이오 연료를 최적화하려고했습니다. . 우리 대부분에게는 e. coli 식품 오염 뉴스 보고서에서만 언급되어 있습니다. 유전자 공학 세계에서 E. Coli는 잘 알려진 성장 대사와 가장 큰 유전자 공학 도구 세트를 가진 귀중한 사용자 친화적 호스트입니다. 많은 사람들 이이 박테리아가 에 대해 광범위하게 연구되었다는 것을 알고 놀랄 수도 있습니다. 유전자 조절 및 발현, 당뇨병 치료를위한 인슐린 생산과 같은 인간 사용을위한 귀중한 물질의 공급원이되었습니다. 대장균 콜라이 다양한 설탕을 섭취하여 실험실 및 산업 규모 모두에서 다재다능한 성장 조건을 허용하여 e를 만듭니다. coli 유전자 공학을위한 선택의 유기체.
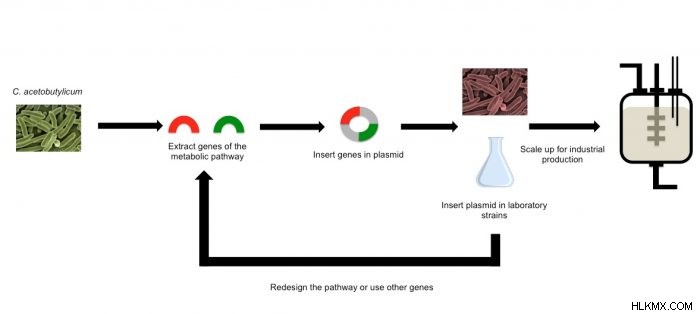
유전자 공학의 발전은 유전자가 이소프로판올과 같은 특정 잠재적 바이오 연료를 생산하도록 유전자를 변형시킬 수있게 해주었다. 이러한 변형은 새로운 유전자를 도입하거나 기존 유전자를 변형시킴으로써 유기체의 유전자 구성을 변화시킨다. 이것은 DNA 염기 쌍 (아데닌-티민/구아닌-사이토신)을 변경하거나 전체 유전자를 삭제하거나 여러 유전자를 삽입하여 수행 할 수 있습니다. 생물학적 엔지니어가 한 유기체에서 다른 유기체로 전체 대사 경로를 전달할 수있는 도구로 인해 과학자들은 실험실에서보다 쉽게 설계된 유기체와 협력 할 수 있습니다. 이러한 도구는 일반적으로 바이러스 및 박테리아와 같은 유기체에 사용될 수 있으며, 유전자 또는 유전자 서열을 다른 유기체로 옮기면이 유기체가 이소프로 파놀과 같은 관심 분자를 생산할 수 있습니다.
아세톤과 이소프로판올은 지속 가능하고 저렴하며 에너지 밀도가 높으며 대체 연료로 테스트되었습니다. 이 바이오 연료는 이미 기존 엔진과 호환되며 (Poh &Poh, 2017), 이소프로판올은 부식 효과가 줄어든 에탄올보다 더 높은 블렌딩 기능을 갖습니다. 이소프로판올과 마찬가지로 아세톤은 에탄올에 비해 에너지 밀도가 높고 혼합 용량을 가지며 엔진에 부식성이 적습니다. 이러한 모든 특성은 이소프로판올과 아세톤이 바이오 연료 시장에서 에탄올을 보완하거나 대체 할 가능성이 있음을 의미하며, 이들 모두는 clostridium acetobutylicum이라는 박테리아에 의해 자연적으로 생성된다.
Clostridium acetobutylicum 단일 대사 경로를 통해 이소프로 파놀 및 아세톤을 생성하는데,이 유기체는 동일한 서열의 유전자를 사용하여 이들 잠재적 바이오 연료를 생성한다는 것을 의미한다. 아세톤은이 유전자 경로의 주요 생성물로서 생성되며, 이소프로판올은 아세톤으로부터 부산물로서 얻어진다. 이 경로는이 두 분자의 생산을 최대화하도록 설계되지 않았지만 그렇게하도록 설계 될 수 있습니다. 불행히도, c와 함께 사용할 수있는 유전자 공학 도구. 아세토 부틸 리움 바이오 연료 수확량을 최적화하기 위해 제한적이며 다른 몇 가지 생화학 적 경로를 억제해야합니다 (Toogood &Scrutton, 2018).
다행히도,
유전자 공학의 발전으로 과학자들은 인간이 건강, 에너지 및 음식 이점을 위해 대량 생산할 수있는 화합물을 생산하기위한 유기체의 자연 대사 경로를 연구하고 개선 할 수있게 해주었다. C. acetobutylicum에서 전체 대사 경로를 전달하는 능력 대장균으로 전통적인 연료를 개선하거나 대체 할 수있는 효율적인 고품질 바이오 연료를 생산하여 에탄올과 관련된 신흥 공급 및 환경 문제를 완화시킬 수있는 효율적이고 고품질의 바이오 연료를 생산하기 위해 유기체의 자연 능력을 상업적으로 실행 가능한 적용으로 이어질 수 있습니다. 배출량을 줄이고 에너지 수율을 향상시키기 위해 엔진 및 연료 혼합물 최적화로 여전히 문제가 남아 있지만, 현재 결과는 석유에 대한 의존도를 줄이려는 사명에서 격려하고 있습니다.
.이러한 결과는 Bredesen Center의 Jessica Velez가 Laval University의 통합 및 시스템 생물학 연구소의 Knoxville, Angel Fernando Cisneros Caballero의 Tennessee University, Knoxville University의 학제 간 연구를 위해 수행 한 작업 및 The Immunity, Infection and Respiratory Medicine의 Narjes Alfuraiji에서 실시한 작업을 기반으로합니다.
참조 :
- Stephanopoulos G., (2007). 바이오 연료 생산을위한 미생물 공학의 과제, Science , 315 (5813), 801–804.
- Clemente, J. 바이오 연료가 오일을 대체 할 수없는 이유. 포브스. 2015. https://www.forbes.com/sites/judeclemente/2015/06/17/why-biofuels-cant-replace-oil/#467b62f6f60f
- Poh, C., &Poh, C., (2017). 소규모 무인 항공기의 연료로서 이소프로 파놀, 항공 우주 과학 기술의 발전 , 2 , 23-30.
- Atsumi, S., &Liao, J., (2008). 대장균의 고급 바이오 연료 생산을위한 대사 공학 , 생명 공학의 현재 의견
- , 19 (5), 414-419.
- Koppolu, V., &Vasigala, V., (2016). 대장균의 역할 바이오 연료 생산에서 미생물학 통찰력 , 9 , 29-35.
- Toogood, H., &Scrutton, N., (2018). 알코올의 발효 생산을위한 미생물 재고, 생명 공학의 현재 의견 , 50 , 1-10.