중심체는 세포 내 소기관으로서 세포의 미세 소관을 구성하는 일차적 인 기능이다. 미세 소관은 세포의 골격으로 생각 될 수 있으므로 중심체는 세포 골격을 구성하고 고정시키는 린치 핀 및 스캐 폴딩 역할을하는 소기관입니다.
센트로 좀은 또한 세포가 두 개의 동일한 사본으로 분할하여 세포가 재현하는 과정 인 세포 유사 분열에서 역할을합니다. 중심체는 염색체에 부착 된 구조 인 유사 분열 스핀들을 형성하는 데 도움이됩니다. 센트로 좀은 1883 년 에두 오드 반 베네덴 (Eduoard van Beneden)에 의해 처음 발견되었으며 1888 년에 공식 이름을 받았습니다.
중심체는 진핵 생물 중종 (다세포 동물)에서만 발견됩니다. 진핵 생물 유기체에서 센트로 좀은 m 역할을합니다 icro t ubule o c 셀의 (MTOC)를 입력하십시오. 식물과 곰팡이는 다른 MTOC 구조를 사용하여 미세 소관을 구성합니다. 다세포 동물에서, 센트로 좀은 세포가 잘 정의 된 모양과 구조를 갖는 주된 이유 중 하나입니다.
중심체는 효율적인 방사형 패턴으로 미세 소관을 구성하여 세포의 모양을 유지합니다. 센트로 솜은 또한 세포 분열을 시작하는 중심을 제공하여 세포 유사 분열 동안 복사의 오류를 방지합니다. 중심 조직 구조의 부족으로 인해 세포가 유사 분열을 시작하기 위해 적합한 국소화 부위를 찾는 것을 방지하기 때문에 중심체가없는 세포는 유사 분열에서 덜 효율적 인 경향이있다.
.센트로 좀의 조성/구조
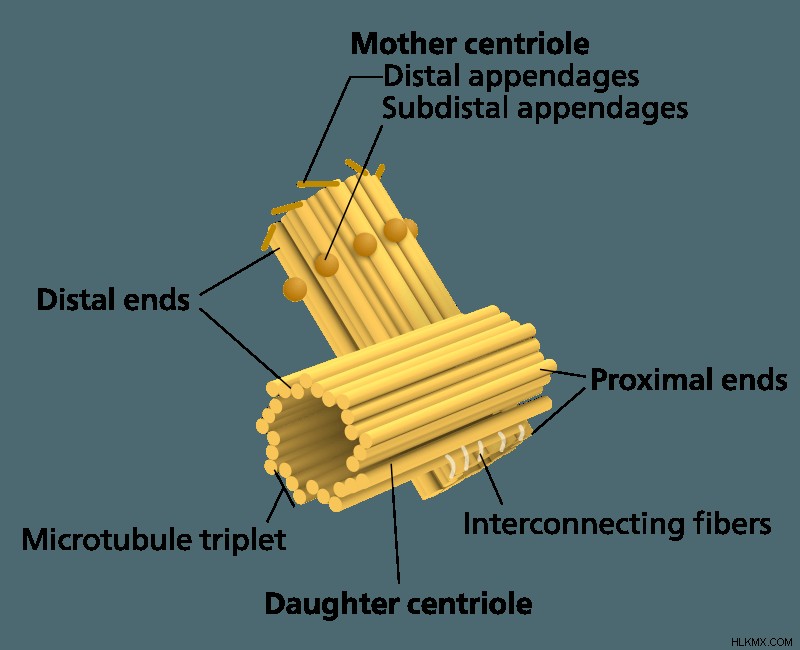
센트로 좀은 centrioles라는 2 개의 서브 유닛으로 구성된 소기관 복합체입니다. 서로 수직으로 배열. 복합체를 둘러싼 것은 pericentriolar 물질이라고 불리는 다양한 단백질의 메쉬입니다. 주변 중심 물질은 미세 소관 어셈블리 및 고정을 담당하는 단백질을 함유하고; γ- 튜 불린, 페리 텐 린 및 9in. 페리 센트리올라 물질은 센트리올을 함께 유지하고 미세 소관의 결합 부위 역할을합니다.
각각의 중심선 자체는 튜브 형 모양으로 배열 된 일련의 미세 소관으로 구성됩니다. 미세 소관은 α- 튜 불린 및 β- 튜 불린의 두 종류의 단백질로 구성된다. 미세 소관은 α- 튜 불린의 이량 체 및 β- 튜 불린이 편광 된 사슬에 동의 할 때 형성됩니다. 이 편광 사슬은 단일 방사형 축 주위에 원형 패턴으로 배열됩니다. 가장 일반적으로, 각각의 중심은 27 개의 개별 미세 소관으로 구성되며, 3의 그룹으로 방사형으로 배열되어있다. 예를 들어, 과일 파리에는 트리플 가닥 대신 이중 가닥의 미세 소관으로 구성된 중심이 있습니다.
각각의 중심체에는 어머니라고 불리는 2 개의 중심이 포함되어 있습니다 및 딸 중심. 어머니 센트리올은 더 오래된 쌍 중 하나입니다. 세포 분열에 앞서 DNA 복제 동안, 기존 어머니와 딸 센트리 올리올의 근위 끝에서 2 개의 새로운 중심이 형성 될 것이다. 새로 구성된이 중심은 각각 새로운 쌍의 딸이됩니다. 유사 분열 동안, 원래의 어머니-딸 쌍은 세포 분열로 분리되어 유사 분열 동안 형성된 각각의 새로운 세포에 모자 딸 쌍을 제공합니다. 어머니와 딸 중심은 각 중심의 근위 끝에있는 섬유와 함께 묶여 있습니다.
다른 미세 소관 구조와 비교하여, 중심체는 매우 강력하고 안정적입니다. 대부분의 미세 소관은 다소 빠르게 분해되므로 세포가 끊임없이 생산하기 위해 노력하고 있습니다. 반면에 중심 미세 소관은 장기 안정성에 기여하는 특수 중심 -특이 적 단백질과 함께 유지됩니다. 또한 이러한 증가 된 안정성은 부분적으로 센트리올 미세 소관의 건설 후 변형으로 인한 것으로 생각된다. 길이 조절 메커니즘에 대한 심층적 인 이해는 아직 없지만 중심 길이를 조절하는 메커니즘이있는 것으로 보입니다.
센트로 좀의 기능
중심체의 주요 기능은 세포에서 미세 소관의 분포 및 배향의 생성을 자극하고 구성하는 것입니다. 미세 소관은 세포에서 모양/구조를 제공하는 것에서부터 세포 내 수송을위한 일종의 "고속도로"역할을하는 것부터 많은 기능을 수행합니다. 따라서, 미세 소관을 생성하고 구성하는 데 중심의 작용은 세포가 작동하는 물리적 프레임 워크를 제공함으로써 거의 모든 다른 셀 작동에 영향을 미칩니다.
.중심체는 미세 소관의 고유 극성 덕분에 미세 소관을 공간적으로 배향 할 수있다. 각각의 미세 소관에는 + 끝 및 A - 끝이 있으며, 이는 다른 세포 구조와 상호 작용하는 방법을 결정합니다. - 미세 소관의 끝은 중심체로 끌려 가므로 + 끝이 세포의 외부를 향해 향하게됩니다. 연장에 의한 중심 및 중심체는 미세 소관 핵 생성을 담당하기 때문에, 이들은 미세 소관의 위치 및 방향을 결정할 수있다. 페리 센트리 올 물질의 단백질은 튜 불린 단백질의 생성을 자극한다. Pericentriolar 물질에 위치한 γ- 튜 불린은 + 편광 끝을 모방하여 고리 형 복합체로 배열되어-미세 소관의 끝을 결합 할 부위를 제공합니다.
중심체는 세포 생식에서 중요한 역할을합니다. 중심체는 유사 분열 스핀들의 핵심을 형성합니다. DNA 복제 동안, 세포 분열의 첫 번째 단계 인 중심체 복합체가 복사된다. 형성 후, 2 개의 중심체 (각각 2 개의 중심체)는 세포의 반대쪽 끝으로 이동하여 유사 분열 스핀들을 구성하기 시작한다. 이 시점에서, 세포의 핵을 둘러싼 막이 용해되어 염색체를 노출시킨다.
스핀들은 주로 키네토 코레 (kinetochores)라는 특수 단백질의 작용을 통해 세포의 염색체와 연결되는 미세 소관으로 구성됩니다. 세포의 두 극에있는 2 개의 중심체의 위치는 두 염색체를 서로 위에 정렬시켰다. 세포 염색체의 이러한 재정렬은 정확한 염색체 분열과 세포 분열의 평면을 지정하기 위해 필요합니다. 분열 평면을 나타내는 셀을 통과하는 중심선을 Metaphase 플레이트라고합니다.
스핀들 장치는 염색체에 부착 된 + 끝을 갖고 세포의 극에 위치한 중심체에 부착 된 끝이 있으며, 세포의 실제 분할이 발생합니다. 스핀들 장치의 중앙 섹션에서, 미세 소관은 해수 중합되어, 두 극 방향의 방향으로 수축하고 장력을 발휘할 수있다. 이어서, 부착 된 염색체는 분리되어 각각 한 쌍의 염색체를 갖는 두 개의 별개의 핵으로 재구성된다. 마지막으로, 세포 세포질은 두 개의 동일한 세포를 나누고 생성합니다.
유사 분열 과정에 크게 관여하지만, 세포 분열에는 센트로 좀이 필요하지 않은 것으로 보인다. 앞에서 언급 한 바와 같이, 식물과 곰팡이 세포에는 중심체가 없지만 여전히 유사 분열이 있습니다. 식물 및 곰팡이 세포에서, 다른 세포 구조는 미세 소관 조직 센터로서 역할을 수행한다. 진핵 생물에서도 중심체는 유사 분열에 필요하지 않은 것 같습니다.
일반적인 과일 파리 drosophila 실험 센트로 솜이 장애가있는 경우에도 세포 분열이 발생할 수 있음을 보여줍니다. 따라서 Centrosome의 주요 목적은 세포 분열을 가능하게하는 것이 아니라 훨씬 더 효율적이고 정확하게 만드는 것이라고 생각됩니다. 센트로 솜이 부족한 세포는 잘못 복제 할 가능성이 더 높아서 조기 세포 사멸, 잘못된 물리적 발달 또는 잠재적으로 유해한 유전자 돌연변이를 초래합니다. 예를 들어 과일 파리는 유충에서 중심체가없는 성인으로 발병 할 수 있습니다. 발달하는 동안, 세포는 중심체의 작용없이 정상적으로 자신을 배열합니다. 그러나, 그러한 방식으로 발달하는 과일 파리는 성인에 도달 한 직후에 죽을 것입니다.
센트로 좀 수차 및 암
중심체 구조 및 기능의 수차 및 결함은 다양한 암 및 종양 성장과 관련이 있습니다. 암 세포는 과량의 심막 물질로 인해 너무 큰 중심체를 갖는 경향이 있습니다. 또한, 종양은 종종 부적절한 양의 센트로 좀을 갖는 세포를 포함합니다. 잘못된 수의 센트로 좀의 기원은 부적절한 DNA 복제로 인한 게놈 불안정성과 관련된 다수의 잠재적 메커니즘을 가지고 있습니다.
1902 년, 독일 생물 학자 Theodor Boveri는 세포에서 중심체의 수가 증가한 것으로 가설을 세웠다. 그는 세포를 나누는 중심체의 과도한 활성이 염색체의 완전성을 손상시켜 게놈 불안정성과 악성 종양의 형성을 초래한다고 추론했다. 그의 원래 이론은 여러 측면에서 정확했지만,이 시점에서 중심체 증폭과 암 사이의 정확한 인과 관계는 알려져 있지 않습니다. 중심체 수차는 암 종양의 진행을위한 일종의 피드백 메커니즘을 제공 할 가능성이있다. 수차 중심체의 존재는 부적절한 세포 분열에 기여하며, 이는 수차 센트로 솜 등을 갖는 더 많은 종양 세포로 이어진다.
.센트로 좀의 진화
중심체는 여러 초기의 진핵 종에 존재하기 때문에 진화론 적으로 오래된 소기관이다. 센트로 좀에 대한 코딩되는 유전자는 센트 린이라고하며 모든 진핵 생물 유기체의 초기 공통 조상에 존재합니다. 대조적으로, 고풍과 박테리아에는 센트 린과 유사한 유전자가 없으므로 중심의 존재는 진핵 생물 유기체의 독특한 특징을 구성한다. 과일 파리와 같은 종은 비정상적인 중심체 구조를 설명하는 두 가지 주요 중심 그룹 중 하나를 잃었습니다.