자연에는 세포와 공생 박테리아, 또는 내부에서 살고있는 내부 대증 사이의 관계가 거의 없습니다. 이러한 파트너십에서, 숙주 세포는 일반적으로 내생형에 대한 보호 기능을 제공하고 전파하는 방법을 제공하는 반면, 내생형은 숙주에 중요한 영양소를 제공한다. 그것은 숙주와 내배엽의 게놈이 서로의 대사 및 생합성 경로에 보완적인 부분을 기여하는 것처럼 보이는 깊은 협력적인 배열입니다.
이러한 파트너십의 공개 된 복잡성은 계속 놀라운 일을하고 있습니다. 오늘날 Cell 에 나타나는 새로운 연구에서 과학자들은 곤충 세포와 두 종의 박테리아 사이의 복잡한 3 방향 공생 (하나는 다른 하나의 내 endosymbiont)이 유기체의 게놈과 생리학과 깊이 얽혀 있음을 보여줍니다. 이러한 결과는 가장 빠른 진핵 세포의 고대 내각에서 미토콘드리아와 다른 소기관이 어떻게 생겼는지를 밝힐 수 있습니다.
세포가 생존하기 위해 새로운 대사 특성을 신속하게 얻어야 할 때, 그들의 최선의 선택은 다른 유기체에서 하나를 빌리는 것일 수 있습니다. 수평 전이는 세포들 사이에 몇 가지 유전자를 움직일 수 있지만, 복잡한 대사 경로를 위해 수평 적으로 유전자 첨가를 획득 할 수있는 가능성은 사라지게 작다. 따라서 가장 쉬운 솔루션은 종종 몬태나 대학교의 내생화 연구원 인 존 맥 슈치온 (John McCutcheon)에 대해 설명합니다. 이 합병은 본질적으로 드문 일이 아닙니다. 이차 및 3 차 합병은 심지어 발생한 것으로 알려져 있으며, 중첩 된 러시아 인형 세트와 동등한 셀룰러를 생성합니다.
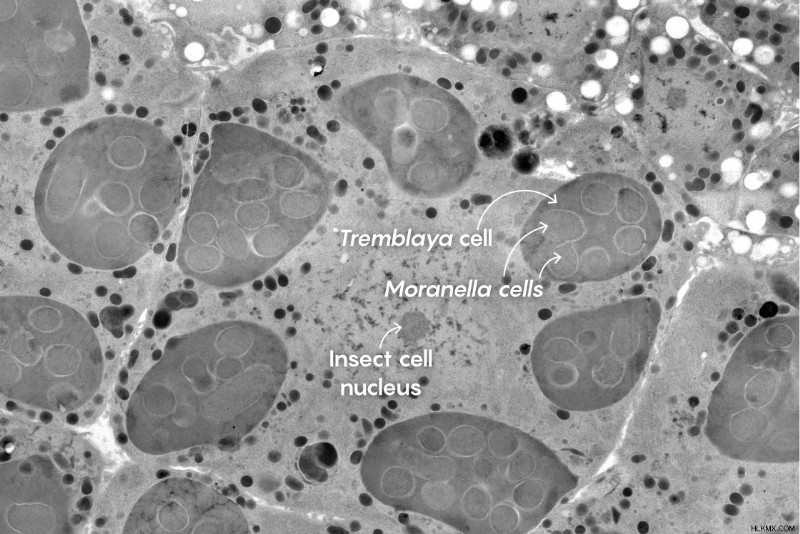
그러한 러시아-홀 합병 중 하나는 약 1 억 년 전에 Mealybugs라는 작은 곤충 해충이 박테리아 내피를 얻었을 때 발생했습니다. . 그 후, tremblaya moranella 을 포함한 다른 여러 박테리아를 획득했습니다 . 결국 다른 사람들은 잃어 버렸고 moranella 만이되었습니다 남아있었습니다. moranella 가 얼마나 오래 걸리는지는 알려져 있지 않습니다 endosymbiosis는 계속되고 있지만 아마도 수천만 년의 순서 일 것입니다. 그 결과, Mealybug Cell에는 2001 년 유타 주립 대학의 생물학자인 Carol von Dohlen이 발견 한 배열 인 또 다른 박테리아가 포함 된 박테리아가 포함되어 있습니다.
.2011 년 Von Dohlen과 McCutcheon 은이 두 박테리아의 서열 분석 된 게놈을 발표했습니다. 각각의 게놈은 유전자를 잃어 버렸지 만, 함께 필수 아미노산을위한 생합성 경로에서 효소를 코딩하는 유전자의 전체 보완을 가졌다. 따라서 tremblaya 및 Moranella Mealybug가 엄격한 SAP 다이어트에서 찾을 수없는 필수 아미노산을 생산하기 위해 함께 일하십시오.
그러나 두 박테리아 게놈은 다른 유전자도 누락되었으며, 아미노산 합성을 위해 서로 보완했지만 효소를 다른 대사 경로에 결정적으로 만들 수없는 것처럼 보였다. 그로 인해 맥 슈치온은 숙주 곤충의 게놈에 구멍을 채운 유전자가 포함되어 있는지 궁금해했습니다.

2013 년에 출판 된 논문에서 McCutcheon과 동료들은 이것이 실제로 그 경우라는 것을 보여주었습니다. 그들은 또한 그 유전자가 곤충 숙주 세포의 핵 내부에 앉아 있었지만, 많은 이들은 박테리아 세포벽의 주요 성분 인 펩티도 글리 칸 합성을 코딩하기 때문에 mealybug 유전자로 분명히 시작되지 않았다는 것을 알아 차렸다. 이 유전자는 박테리아로부터 수평으로 mealybug 핵 게놈으로 전달되어야했다.
따라서 게놈 증거는 moranella Endosymbiont는 Mealybug의 핵 게놈에서 유전자 생성물에 의존하여 세포벽을 만들 수 있습니다. 그러나 그렇다면 곤충 유전자의 생성물은 5 개의 세포막 (3 인치 tremblaya )을 통해 숙주 핵에서 이동해야한다는 것을 의미했습니다. 그리고 moranella in ) 펩티도 글리 칸이 만들어지는 가장 깊은 중첩 박테리아의 내부에 도달합니다. 그 입증되지 않은 제안은 가능성이 거의 없었습니다.
또한, 적어도 세 가지 이종 공급원의 유전자 (정통 Mealybug 핵 유전자, Mealybug 핵 게놈에 의해 획득 된 다양한 박테리아 유전자, 및 Moranella )를 의미합니다. 유전자 - 모두 복잡한 생합성 경로에서 함께 작동했습니다. 그것은 또한 가능성이 낮은 것처럼 보였습니다.
가설이 형성되는 가설은 심지어 연구원들에게도 틀림없이 보였다. McCutcheon은“너무 복잡 하고이 모든 것들이 함께 일하기 위해서는 거의 말도 안됩니다. 그럼에도 불구하고, 그들은 게놈 데이터에 대한 충분한 확신을 가지고 그것을 테스트하는 방법을 고안했습니다. "우리는 우리가 들어갈 수있는 경로를 알아 내고 실제로 유전체학에서 우리가보고있는 것이 실제로 우리가 생각했던 방식으로 작동했음을 증명하고 싶었습니다."
.
공생 및 신진 대사
Cell 에 오늘 출판 된 논문에서 , McCutcheon, Deanna Bublitz (McCutcheon의 실험실의 선임 과학자)와 동료들은이를 달성 할 수있는 영리한 트릭을 설명합니다. 그들은 펩티도 글리 칸의 독특한 특징을 이용했다. 그들은 세포 대사에서 다른 곳에서 발견되지 않은 아미노산 인 D- 알라닌으로 만들어졌다. 실험에서, 연구자들은 D-Alanine의 Mealybug 세포 버전의 배양 물이 무거운 질소 동위 원소 또는 형광 화합물로 태그를 지정하여 세포의 위치와 다양한 대사 변형을 추적 할 수있게 해주었다.
.그들의 연구 결과는 펩티도 글리 칸 합성의 완전한 생화학이 중첩 된 moranella 내에서 발생하고 있음을 확인했다. endosymbionts.
연구원들이 처음으로 중위 동위 원소 실험 결과를 보았을 때, Bublitz는 회상했다. "화면이 나오고 Peptidoglycan으로 우리가 예상 한 패턴이 실제임을 보여주기 시작했지만, 복도에 서서 우리 가보고있는 것이 여전히 눈에 띄는 지 확인하기 위해 매우 멀리 떨어진 화면을보기 위해 잠시 시간이 걸렸습니다."라고 그녀는 말했습니다. "그 수준의 불신이었습니다." 연구원들은 심지어 낯선 사람들에게 전화를 걸어 화면의 이미지를 설명하기 위해“우리와 마찬가지로 맹인 리뷰어만큼 분명하다면, 우리가보고 싶은 것을보고 있지 않았다”고 말했다.
McCutcheon은“저를 가장 흥분하게 만드는 것은이 복잡한 엔도 족이 작동한다는 것을 보여줍니다. 그는 또한 moranella 에 강타했다 및 tremblaya Mealybug 세포에 너무 통합되어 효과적으로 그들 중 일부가 될 수 있습니다.“기초가되는 유전 적 복잡성을 보는 것은 실제로 내생과 소기관 사이의 모든 기능적 구별을 침식합니다.”
.Nova Scotia의 Dalhousie University의 진화 및 분자 생물 학자 인 W. Ford Doolittle 은이 연구는 게놈 데이터에 의해 암시 된 생화학을 테스트했으며 실제로 세포가 그들이 만들고자하는 것을 만들고 있음을 보여 주었기 때문에“필요하고 흥미로운 지상 트 루싱”을 나타냅니다. "나는 이것이 매우 중요한 논문이라고 생각한다"고 그는 말했다.
샌프란시스코 캘리포니아 대학교 (University of California)의 동물과 미생물 사이의 상호 작용을 연구하는 생화학자인 Seekay Chou는“실제로 가장 안쪽 박테리아가 세포벽을 만들어 낸다는 것을 보여줄 수있는 것은 매우 투어 드 포스입니다. "기술적으로는 매우 어려운 일입니다."
“이것에 대해 매우 거친 것은 노동의 분열이 숙주가 5 개의 다른 지질 막을 가로 질러 내부 내부 상징에 도착하기 위해 무언가를 운반해야한다는 것입니다.”라고 그녀는 덧붙였습니다.
이 시점에서 공생 세포가 진핵 생물 세포가 세포막을 가로 질러 단백질을 셔틀에 사용하는 데 일반적으로 사용하는 것과 동일한 분자 메커니즘을 사용하고 있는지 또는이화물을 움직일 수있는 새로운 방법을 발명 해야하는지는 확실하지 않습니다. McCutcheon에 따르면, 일부 증거는 tremblaya 을 나타냅니다 ,이 3 방향 공생의 중개인은 유전자 생성물을 숙주 핵과 Moranella 사이의 적극적으로 운반합니다. 게놈, 무거운 동위 원소 태그 대사 산물이 tremblaya에서 끝나지 않았기 때문에 . "어떻게 든 참여하고 있지만‘어떻게’는 정말 미스터리입니다."
공유 유전자를 포함하는 내성 상징적 인의 다른 예제가 하나 이상 있습니다. Paulinella , 프로테이스트는 시아 노 박테리아 내피온을 획득함으로써 약 1 억 년 전에“두 번째 엽록체”또는 크로마토 포어를 진화시켰다. 크로마토 포어 게놈은 크로마토 포어의 펩티도 글리 칸 층을 만들기 위해 숙주 프로테이스트 게놈을 보완하는데 작용한다.
McCutcheon은 Paulinella 모두에서 추측했다 그리고 Mealybug- tremblaya-moranella 펩티도 글리 칸을 만들기위한 게놈 모자이크 인 endosymbiosis는 진핵 생물 숙주가 박테리아 내배열을 제어 할 수있게한다. 엔도 족이 너무 빨리 복제되면 호스트를 죽일 수 있습니다. 박테리아가 세포벽을 만들 수있는 속도를 제한함으로써 호스트는 거주자를 점검하게합니다. 왜 moranella 그러나 세포벽은 계속해서 미스터리로 남아 있습니다. 그리고 호스트 셀. Chou는“분명히이 모든 일을해야한다는 사실은 어떤 이유로 든 중요하다는 것을 암시합니다.
McCutcheon은 Mealybug ménage à trois가 매우 오래되고 아마도 가장 잘 알려진 오르코네의 진화에 대한 단서를 가질 수 있다고 생각합니다.
미토콘드리아는 15 억에서 20 억에서 20 억 사이의 원핵자 (아마도 Archaea의 구성원)에 의해 휩싸인 알파 프로 테오 박테리아에서 진화했습니다. 대부분의 살아있는 박테리아에는 펩티도 글리 칸 세포벽이 있기 때문에 미토콘드리아의 조상을 포함한 고대는 그랬을 것입니다. Bublitz는 알파 프로 테오 박테리아가 다른 세포에 의해 취해 지거나 적극적으로 침입했으며, 숙주 세포는 펩티도 글리 칸 경로를 조합하여 내생형의 복제를 제어 할 수 있다고 가정합니다. 호스트와 초기 미토콘드리아가 결국 더 통합 될 가능성이 높아서 호스트는 더 이상 미토콘드리아의 분할을 제어하기 위해 펩티도 글리 칸 경로가 필요하지 않습니다. 펩티도 글리 칸 합성에 대한 제어 제어는“자율 박테리아에서 어떤 종류의 기능성 소기관으로 전환하는 첫 단계 중 하나 일 수있다”고 Bublitz는 추측했다.
미토콘드리아는 너무 고대이기 때문에 한 번만 진화했기 때문에 그 내피증이 어떻게 진화했는지 정확하게 재구성하기가 어렵습니다. 우리가 아는 것은 시간이 지남에 따라 미토콘드리아 게놈이 유전자를 잃어 버렸다는 것입니다. 그 중 일부는 핵 게놈에 삽입되었습니다. 오늘날, 대부분의 동물에서 양측 대칭을 가질만큼 복잡한 미토콘드리아 게놈은 37 개의 유전자 만 유지합니다. 미토콘드리아는 현재 핵에서 1,000 개가 넘는 유전자에 의존합니다. (대조적으로, 진핵 생물 미생물은 일반적으로 3 내지 69 개의 미토콘드리아 유전자를 가지고있다.) 그러나 그 핵 유전자 중 다수는 고대 미토콘드리아 게놈에서 나오지 않았다는 사실이다. 그들은 수평 전이 사건에서 다른 박테리아에서 나왔다고 McCutcheon은 말한다.
이 수평 전이 된 유전자의 기원은 미토콘드리아 생물학에서 뜨거운 논쟁의 여지가있다. 한 가지 가능성은 진핵 세포를 생성하는 내 세포 상징적 사건 전에 알파 프로 테오 박테리아가 다른 박테리아로부터 획득했을 것이며, 그 후 유전자는 이후에 숙주 핵으로 이동했다. 다른 하나는 수평 전이 유전자가 진핵 세포가 진화 한 후 시간이 지남에 따라 다양한 박테리아로부터 직접 핵 게놈에 도착했다는 것이다.
우리는 가장 오래된 소기관이 어떻게 진화했는지 완전히 확신 할 수는 없지만 Mealybug- Tremblaya-Moranella 공생은 두 번째 진화 시나리오가 효과가 있음을 보여줍니다. 다른 박테리아 감염의 유전자가 천천히 축적되어 단일 실체에서 기능적 경로로 통합 될 수 있음을 보여줍니다.
McCutcheon은 McCutcheon은 Endosymbionts-Turn-organelles에서 Mitochondria를 독특하게 만드는 것은 가장 오래된 예라고 말했다. “그러나 그들의 유물은 공부하기가 어렵고, 소기관이되기 전에 일어난 일을 추론하기가 어렵습니다. 나는 paulinella 에 관한 것이라고 생각합니다 그것은 우리에게 일어날 일에 대한 창을 제공한다는 것입니다.”
미토콘드리아가 독특하게 성공했지만 모든 내생형이 행복하게 끝나는 것은 아닙니다. McCutcheon의 초기 연구는 시카다스가 박테리아 내 endosymbiont, hodgkinia cicadicola 을 가지고 있음을 보여주었습니다. , 그것은 Cicada 세포 내부에 24 개 이상의 계통으로 조각화되었습니다. 이 계통은 각각 hodgkinia 의 다양한 서브 세트 만 포함합니다. 게놈. 함께, 모든 계통은 시카다가 의존하는 필수 아미노산을 만드는 데 필요한 유전자를 완전히 보완합니다. 그러나 McCutcheon은 이것이 호스트에게 어려움을 야기하는 비 적응성 내생화 진화의 예라고 생각합니다. Cicada Eggs는 생존을 위해 내생형의 완전한 보완 물을 선택해야하며, 엔도 족의 다른 조합이 포즈 문제를 제공하는 것에 대한 변동성이 필요합니다.
McCutcheon과 Bublitz는 이제 일부 엔도 징시가 왜 안정되고 성공적인 파트너십으로 진화하는 반면, 다른 엔도 족이 통제 불능 또는 저하로 진화하는 이유를 알아 내기 위해 노력하고 있습니다. Bublitz는“지금 당장 흡연 건이 지속될 수있는 것에 대한 흡연 건이 없다”고 Bublitz는 말했다. 통제 파트너