핵심 개념
이 기사에서는 각 DNA 타우토머의 구조 및 생화학 적 특성에 대해 배웁니다 :B-DNA, A-DNA 및 Z-DNA.
.wautomer 란 무엇입니까?
DNA를 생각할 때 친숙한 이미지가 즉시 비틀림 사다리와 같은 이중 나선을 떠올리게됩니다. 그러나, 다른 많은 유기 분자와 마찬가지로, DNA는 특정 환경 조건을 고려할 때 팽창 제라고 불리는 여러 가지 형태를 취할 수 있습니다. 생화학 자에게 B-DNA, A-DNA 및 Z-DNA로 알려진이 DNA 이성질체는 생물학적 시스템에 중요한 화학적 특성을 가지고 있습니다.

화학자들은 두 분자 구조가 자발적으로 서로 전환되는 반응으로서“타우토머 화”를 정의합니다. 팽팽한 이성질체라고 불리는이 두 분자는 헌법 이성질체 인 경향이 있으며, 이는 동일한 화학적 공식이지만 원자 사이의 다른 연결성을 갖는다는 것을 의미합니다. 실제로, 많은 팽팽한 이민자는 수소 원자의 배치 만 다릅니다. 또한, 타우토머는 동적 평형 상태에 존재하는 경향이있다. 이것은 타우토머 A 로의 팽팽한 이민자 B 로의 전환율이 A와 B의 상대적 농도가 시간이 지남에 따라 일정하게 유지되도록 B의 전환율과 같음을 의미한다.
.케토 에놀 호변 이성질체는 유기 및 생화학에서 타우토머 쌍의 가장 일반적인 예를 제공합니다.
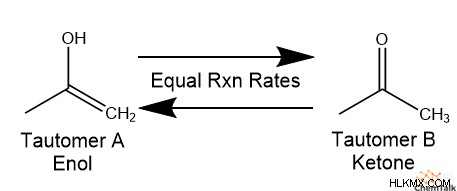
다른 팽팽함과 달리, B-DNA, A-DNA 및 Z-DNA는 분자 구조가 다르지 않고 오히려 두 나선 사이의 분자간 상호 작용에서 다릅니다.
B-DNA 호텔 이머
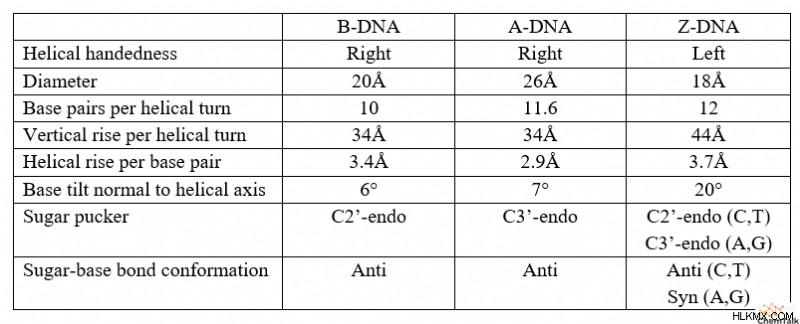
과학자와 비 과학자 모두에게 B-DNA는 가장 잘 알려진 형태의 DNA 역할을합니다. X- 선 결정학을 사용하여 Rosalind Franklin에 의해 원래 이미지화 된 B-DNA는 DNA의 Watson 및 Crick 모델의 기초로 사용되었습니다. B-DNA는 손, 기본 쌍 위치 및 설탕 주름을 통해 다른 팽팽한 이민자와 다릅니다.
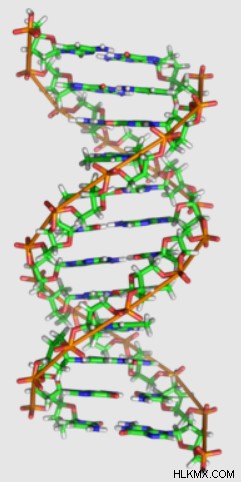
먼저, B-DNA 타우토머는 바람이 불어“오른 손잡이”이중 나선을 형성한다. 이것은 가닥이 시계 방향으로 위쪽으로 비틀어지는 "왼손잡이"이중 나선과는 반대로, 시계 방향으로 위쪽으로 비틀어지는 것을 의미합니다. 이 차이점을 기억하려면 오른손을 당신 앞에서 상상해보십시오. 엄지 손가락으로 위쪽으로 , 손가락을 주먹으로 말리십시오 (또는“엄지 손가락”). 손가락이 카운터 시계 방향으로 을 찾을 수 있습니다 . 왼손과 똑같이하면 Finders가 Curl 시계 방향으로 을 찾을 수 있습니다. 엄지 손가락을 가리키면 위쪽으로 가리킬 때.
둘째, B-DNA의 기본 쌍은 나선 축의 것과 거의 수직 인 평면을 형성한다. 다르게 말하면, B-DNA의 이중 나선을 수직으로 지향하면,베이스 쌍은 DNA 가닥 사이에 거의 완벽하게 수평 브리지를 형성합니다.
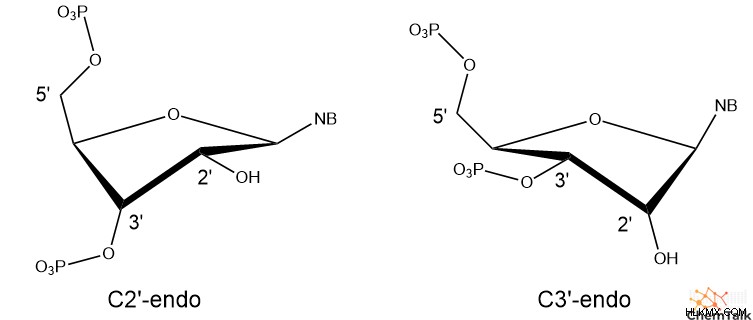
셋째, B-DNA에는 C2'-Endo라는 설탕 주름 형태가 있습니다. 이것은 DNA의 Deoxyribose 설탕의 3D 형태를 나타냅니다. 구체적으로, C2'-Endo Pucker는 설탕의 2 '탄소가 5'탄소에 부착 된 인산염과 동일하다는 것을 나타냅니다. 이 방향은 대체 C3'- 엔도와 비교하여 5 '와 3'포스페이트 사이의 거리를 연장합니다.
a-dna 호텔 이머
B-DNA와 동일한 명성과 인식 가능성이 없지만 Rosalind Franklin의 작업을 통해 A-DNA가 동시에 발견되었습니다.
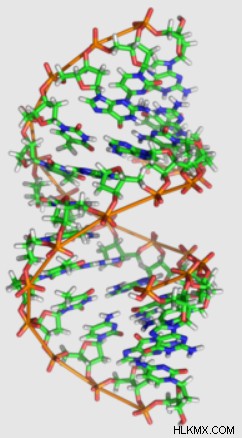
B-DNA와 마찬가지로 A-DNA는 또한 오른 손잡이 나선 구조를 포함합니다. 그러나 A-DNA의 Deoxyribose는 C3'-Endo의 주름을 나타내며, 이는 B-DNA보다 5 '및 3'인산염을 더 가깝게 가져옵니다. 이로 인해 A-DNA는 B-DNA에 비해 짧고 쪼그리고 앉는 모양을 갖습니다.
또한, A-DNA베이스 쌍은 헬리컬 축에 이르기까지 20도 각도로 존재한다. 이 각진베이스 페어링은 또한 B-DNA와 같이 중심을 가로 질러 확장되지 않고 나선의 외부 표면을 따라 가닥을 연결합니다. 이로 인해 이중 나선이 "중공"이되고 구조물의 내부 아래로 밀짚 모양의 구멍이 생깁니다.
생화학 자들은 A-DNA가“탈수 조건”이라고 부르는 것 하에서 형성된다는 것을 발견했다. 이것은 단순히 물 분자가 DNA와 상호 작용한다는 것을 의미합니다. 근처의 물 부족은“소수성 효과”를 약화 시키는데, 이는 비극성 영역이 내부에 차폐되는 동안 생체 분자의 극성 영역이 표면으로 끌리는 현상이다. B-DNA에서, 극성 데 옥시 리보스 및 포스페이트 골격은 나선 외부의 물과 상호 작용한다. 반대로, 질소 염기의 비극성 영역은 B-DNA의 안정성에 중요한 "스태킹 상호 작용"을 형성한다.
A-DNA는 나선 외부를 따라 염기 패링을 포함하기 때문에, 약화 된 소수성 효과만이 A-DNA 타우티머 형성을 가능하게한다.
발견 된 이래, 생화학 자들은 박테리아가 용매를 제거하는 단백질을 사용하여 A-DNA의 형성을 유도 할 수 있음을 발견했다. 아마도 박테리아는 A-DNA 타우토머가 극심한 열 또는 건조 조건으로부터 유전자를 더 잘 보호하기 때문에이를 수행합니다.
흥미롭게도, 이중 가닥 RNA의 구조는 A-DNA의 구조와 유사합니다.
Z-DNA 호텔 이머
Rosalind Franklin이 발견 한 지 수십 년이 지난 후 생화학 자 Andrew Wang과 Alexander Rich는 특정 DNA 서열이 Z-DNA라고 불리는 세 번째 팽팽함을 나타냅니다.
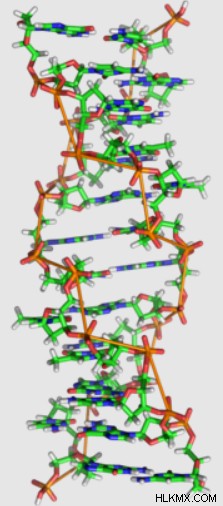
다른 팽팽한 이민자와 달리 Z-DNA는 왼손잡이 나선 구조를 가지고 있습니다. a-DNA 및 B-DNA에 비해 Z-DNA는 더 길고 얇은 헬리컬 구조를 가지고 있습니다.
흥미롭게도, Z-DNA는 퓨린 (아데닌 및 구아닌)과 피리 미딘 (티민 및 시토신)을 교대하는 DNA 서열 사이에만 형성되는 것으로 관찰되었다. 이러한 서열은 몇 가지 다른 서열 중에서 다른 가닥의 구아닌 및 시토신 (폴리 (GC) - 폴리 (GC)) 또는 다른 가닥의 구아닌 및 티민 (폴리) - 폴리 (GT)와 쌍을 이루는 2 개의 가닥을 포함 할 수있다.
Z-DNA에서,이 서열은 흥미로운 구조를 가정하며, 여기서 퓨린은 C3'- 엔도 주름을 가지고 있고 피리 미딘에는 C2'- 엔도 주름이 있습니다. 또한, 설탕과 퓨린 염기 사이의 결합은 기저 고리 구조의 대부분이 데 옥시 리보스 위에 매달린다. A-DNA, B-DNA 및 Z-DNA 피리 미딘에서, 모든 설탕 대 기본 결합은 설탕 반대편에있는 기초가 반응을 보인다.
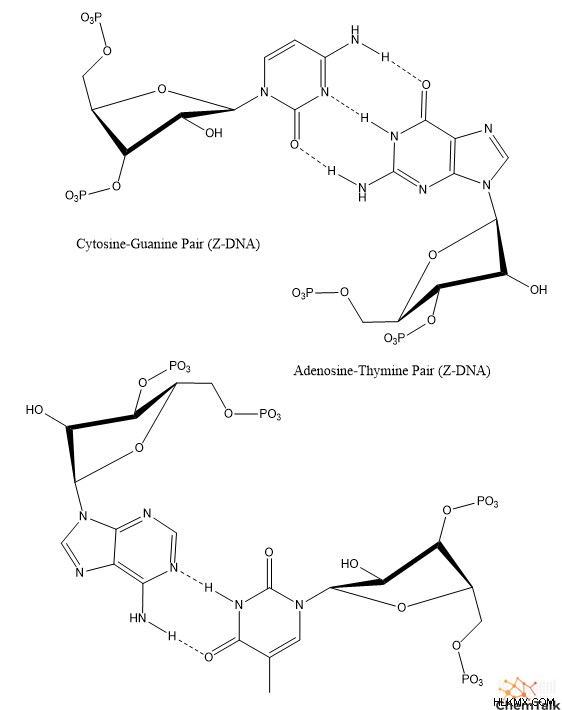
A-DNA와 마찬가지로 Z-DNA는 또한 중요한 생물학적 목적을 가지고 있습니다. 구체적으로, 효소가 전사를 위해 B-DNA를 개방하고 풀어주는 경우, Z-DNA는“음성-수퍼 코일링”을 통해 형성되어 풀링의 스트레스를 완화시킨다. 실제로, 생화학 자들은 Z- 알파 도메인과 같은 Z-DNA와 구체적으로 상호 작용하는 많은 다른 효소 및 결합 단백질을 확인 하였다.
그러나, 서열 사양과는 별도로, Z-DNA는 높은 염 농도 하에서 만 형성되는 것으로 관찰되었다. 생화학 자들은 소금 이온이 Z-DNA에서 훨씬 더 가깝게 하전 된 포스페이트를 안정화시키는 데 도움이된다고 생각합니다. 이러한 이온이 없으면, 음으로 하전 된 포스페이트의 이온 반발로 인해 구조는 안정성을 잃게됩니다.
DNA 타우토머 요약 테이블