생물 다양성 보존은 최근에 다시 언급 한 IPBES (1, 섹션 3.2, pp. 196 참조)로 인류에게 중요한 과제입니다. 최적의 생물 다양성 (2, 3)의 원리는 생물학적 기반으로 돌아와서 자연이 왜 다양성이 필요한지 이해하려고 노력함으로써이를 돕기위한 시도입니다.
다양성이 적응 인 경우
우리는 생물학적 시스템의 내부 다양성 (즉, 구성 요소의 다양성)이 그 효과, 탄력성 및 생존력과 관련된 적응 형 특성이라고 가정합니다. 최적의 다양성을 가진 생물학적 시스템이 가장 효과적이고 실행 가능합니다. 최적의 다양성에서 벗어나는 모든 변화는 생물학적 시스템의 효과와 생존력을 감소 시키거나 결국 그것을 파괴합니다.
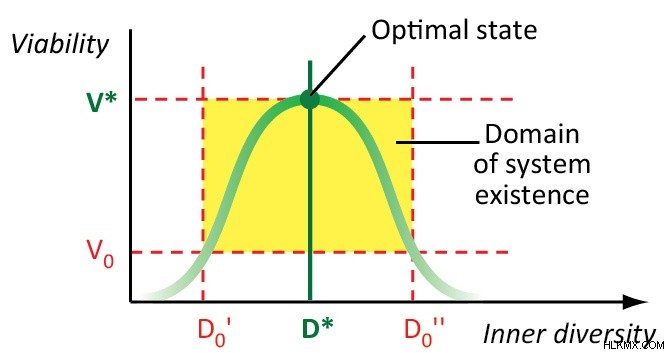
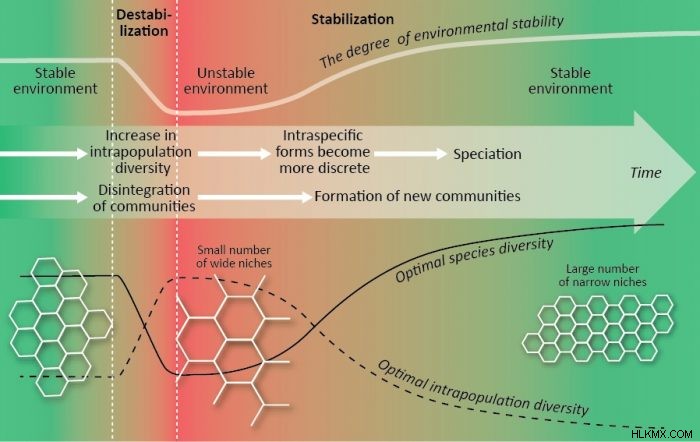
최적의 생물 다양성은 다른 계층 적 수준의 삶의 상호 작용 중에 확립됩니다. 우리의 이론적 모델 (2, 3, 4, 6)은 작은 생명 계층 구조, 즉 인구와 공동체, 즉 인구 구간 및 종 다양성 만 연구했습니다. 추가 사고는이 모델에서 나옵니다.
인구와 지역 사회 수준의 반대 반응 - 같은 동전의 양면
인구 내 및 종 다양성의 최적 값은 반대 모드에서 환경 매개 변수 (이용 가능한 자원의 양과 안정성도)에 따라 다릅니다. 최적의 인구 내 다양성은보다 안정적인 조건에서 감소하는 반면, 최적의 종 풍부도는 증가합니다. 최적의 종의 풍부함은 또한 더 "풍부한"조건에서 증가하며, 최적의 인구 구체 내 다양성은 자원의 양에 의존하지 않습니다. 인구 내 및 종 다양성은 생물학적 계층 구조의 분리 할 수없는 두 가지 측면을 나타내며, 이들의 반응의 차이는 적응 과정에서 특정 역할을 나타냅니다.
상향식 최적화 :구성 요소는 상위 레벨에서 다양성을 정의합니다
최적의 다양성 값은 다음의 모집단 매개 변수에 따라 달라집니다.
- 개인의 생태적 내성 영역의 폭
인구 생존력을 증가시키는 이들 매개 변수의 개선은 환경의 안정화와 같이 최적의 다양성의 동일한 변화를 초래한다. 따라서 인구는 환경 불안정성을 보상하는 다른 방법을 가지고 있습니다. 인구 증가율 증가, 사망률 감소, 개별 허용 범위를 넓히고 표현형의 자원 비용을 낮추십시오. 이러한 메커니즘은 적응 능력으로 인해 인구 내에서 작동 할 수 있으며, 예를 들어 K-와 R- 전략가 사이와 전문가와 일반인 사이의 변화와 같은 종 구성의 변화로 인해 커뮤니티 내에서 작동 할 수 있습니다.
.변하지 않은 남은 다른 사람과의 상기 매개 변수 중 하나의 개선은 진화 수준의 유기체의 증가로 해석 될 수 있으며, 따라서 최적의 인구 내 다양성의 감소와 최적의 종 다양성의 증가를 초래할 수있다.
.최적의 생물 다양성의 예상 패턴
인구 구간의 다양성, 다른 것들이 동등한 것은 생태 틈새의 폭을 대중화하는 것으로 간주 될 수 있습니다. 따라서, 우리는 지역 사회의 최적 생물 다양성 구조를 예측할 수있다 :1) 풍부하고 안정적인 조건에서 좁은 생태 틈새를 가진 다수의 전문가 종, 2) 부족한 불안정한 조건에서 넓은 생태 틈새를 가진 소수의 일반 종, 3) 중간 조건에서 중간 최적 다양성 구조 (6). 방해받지 않은 클라이맥스 커뮤니티와 그 구성 인구는 최적의 다양성에 가장 가깝습니다. 일치하는 생물 다양성 패턴은 전 세계 및 조경 척도 (7)에서 찾을 수 있습니다.
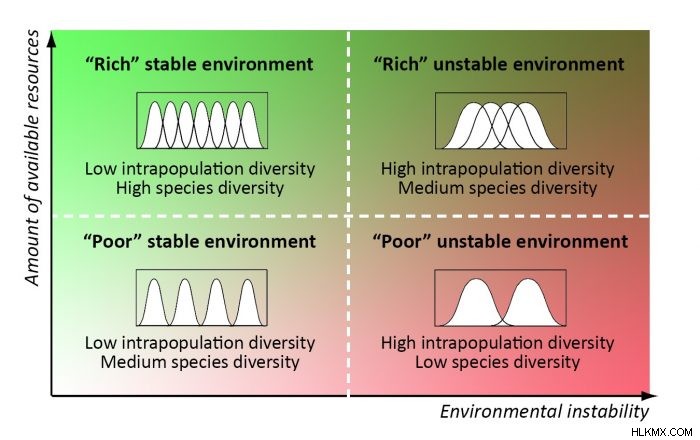
자연 관리 관행의 경우, 이는 보존 우선 순위에 대한 기준이 최적의 다양성에서 멀어지는 인위적인 변화의 거리가되어야하지만, 공동체가 불안정하거나 부족한 상태에 적응 한 공동체는 비교적 낮은 종 다양성을 가지고 있기 때문에 최대 기능을 보장하기 때문에
이기 때문에 공식적인 다양성 지수 (예 :종 풍부함)가 아닙니다.최적의 영원한 추구
바이오 시스템이 최적의 다양성을 달성 할 수 있습니까? 우리의 변형 가능한 우주에서는 거의 볼 수없는 것 같습니다. 환경이나 바이오 시스템에서 자연적 또는 인위적인 변화는 최적의 상태에서 차선책으로 전환합니다. 따라서 생물 시스템은 생태 학적, 미세 조화 및 진화 과정에서 최적의 상태에 대한 영구적 인 "추격"에있다 (동시에 발생할 수 있음)
생태 학적 사례 1 :Synanthropic Biota에서 R- 전략가의 우세.
환경의 인위적 변화의 일반적인 방향은 불안정화이며 (7) 인구에 대한 인간의 영향은 인구 내 다양성의 감소로 표현된다. 후자는 인구가 불안정화에 적응할 가능성을 감소시킨다. 결과적으로, 적응은 공동체 수준에서 작동하기 시작하고 전형적인 원주민 종은 다른 종의 종 구성 작곡으로 대체되며, 종-전략 학자에서 R- 스터디스트로, 전문가로부터 일반인으로 교체되며, 이는 Synanthropic Biota의 확산에 해당합니다.
.생태 학적 사례 2 :커뮤니티 어셈블리의 메커니즘.
커뮤니티 어셈블리에서 특정 메커니즘의 유병률은 환경의“풍부 성”과 안정성 사이의 비율, 또는 오히려 주어진 양의 자원에 존재할 수있는 최적의 틈새의 수와 (예 :t ° C) 변동 (6) :
미시적 진화 예 :다양성 최적화의 메커니즘으로서 Sympatric Intraspecific 형태.
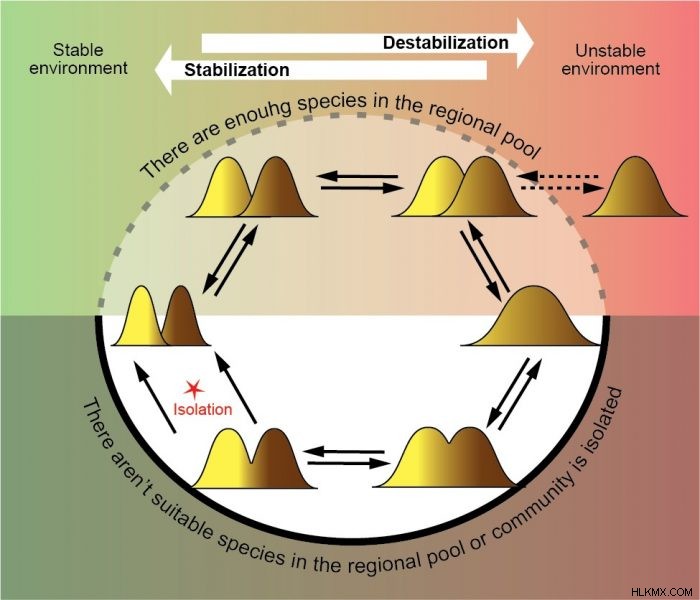
환경 안정성의 정도가 변하는 경우, 공동체는 다른 최적 상태를 이동해야합니다. 안정적인 조건에서 좁은 틈새 시장과 불안정한 조건에서 넓은 틈새를 가진 소수의 종. 지역 수영장에 종이 충분하고 공동체가 최적 성을 달성하기 위해 필요한 특성을 가진 종을 쉽게 포함시킬 수 있다면 문제가 없습니다. 그러나 지역 사회가 고립되어 있거나 지역 수영장에 적합한 종이 없다면 어떻게해야합니까? 이 경우, 한 종이 다중 틈새를 차지할 때 심혈르 내 intraspecific 형태 (생태 타입)의 형성으로 인해 최적화가 발생할 수 있습니다. 이 과정은 특이 적 다양성 (틈새 폭)과 커뮤니티 다양성 (종 수)을 최적화 할 수 있습니다. Sympatric intraspecific 형태는 형태의 분리가 새로운 종의 형성으로 이어질 때까지 지역 생물 다양성을 환경 변화에 지속적으로 적응시키는 역동적 인 시스템으로 작동 할 수 있습니다 (3).
진화론 적 예 :기복에도 불구하고 다양성의 전반적인 증가.
생물 다양성 최적화는 V. Zherihin의 진화에 대한 생체 세게 조절 개념과 V. Krasilov의 일관성 실질 진화 개념 내에서 과정에 대한 추가 설명 일 수 있습니다. 환경 불안정화 및 지역 사회의 파괴 (생물 기생성 위기)의 역사적시기는 인구 구간의 다양성의 증가, 틈새 확장, 특수 종의 멸종, 종 다양성의 감소로 특징 지어집니다. 인구 구간의 다양성의 증가는 미래의 intraspecific 형태 및 speciation의 형성을위한 재료를 제공한다. 환경 안정화와 지역 사회의“새로운 세대”의 역사적 기간은 인구 내 다양성의 감소, 틈새 좁은 틈새, 특이 적 형태의 이산화, 종의 증가, 종 수의 증가로 특징 지어집니다. 진화 수준의 유기체 수준의 증가는 인구 구간의 다양성 (최적의 틈새 시장이 좁아짐)의 감소와 생명의 진화 동안 종 다양성의 일반적인 경향에 대한 추가 설명 일 수있는 최적의 종 다양성의 증가를 초래한다 (3).
참조 :
1) 경쟁 및 니치 분리가 전 세계적으로 최적화 된 경우, 니치 분리가 전 세계적으로“NECHE 분리”의 비율에 따라 결정됩니다. 최적의 "범위"틈새는 대략 동일합니다 (r/r*≈D/d*);
2) 중립 메커니즘은 주로 많은 양의 자원이 주로 환경 매개 변수의 범위보다 훨씬 더 큰 종 수의 존재를 허용하는 매우 "풍부한"환경에서 주로 작동합니다 (r/r*>> d/d*);
3). 자원의 자원은 환경 매개 변수의 범위보다 훨씬 작은 종 수를 허용합니다 (r/r*<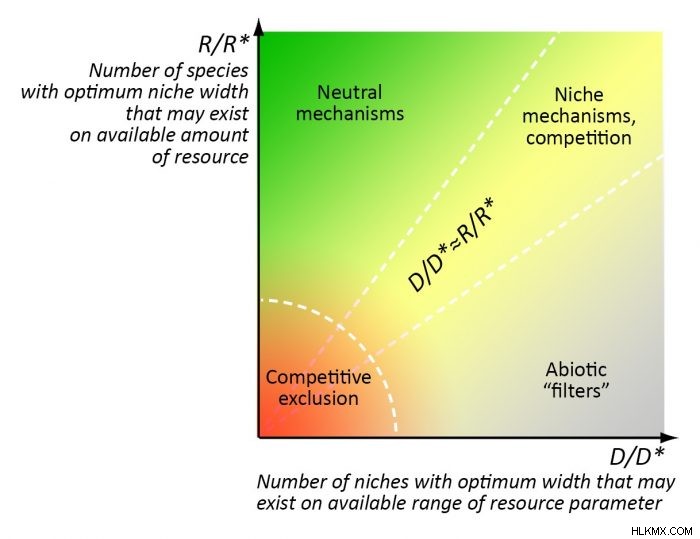
https://ipbes.net/sites/default/files/2018_eca_full_report_book_v5_pages_0.pdf
https://sciencetrends.com/
https://link.springer.com/article/10.1134/s1062359010010012
http://article.sapub.org/10.5923.j.ije.20120204.06.html
http://www.sciencepublishinggroup.com/journal/paperinfo.aspx?journalid=118&doi=10.11648/j.ajls.20130104.16
https://www.sciencedirect.com/science/article/abs/pii/s1470160x17302170?via%3dihub