불완전한 지배력 상속 유형, 특히 특정 특성에 대한 대립 유전자가 쌍을 이루는 대립 유전자에 대해 완전히 (완전히 지배적) 표현되지 않는 중간 상속 유형입니다.
.결과는 발현 된 물리적 특성이 대립 유전자에 속하는 표현형의 조합 인 표현형 (발현)이다. 이 경우 하나의 대립 유전자가 다른 대립 유전자를 가리거나 지배하지 않습니다.
대립 유전자 및 멘델 유전학
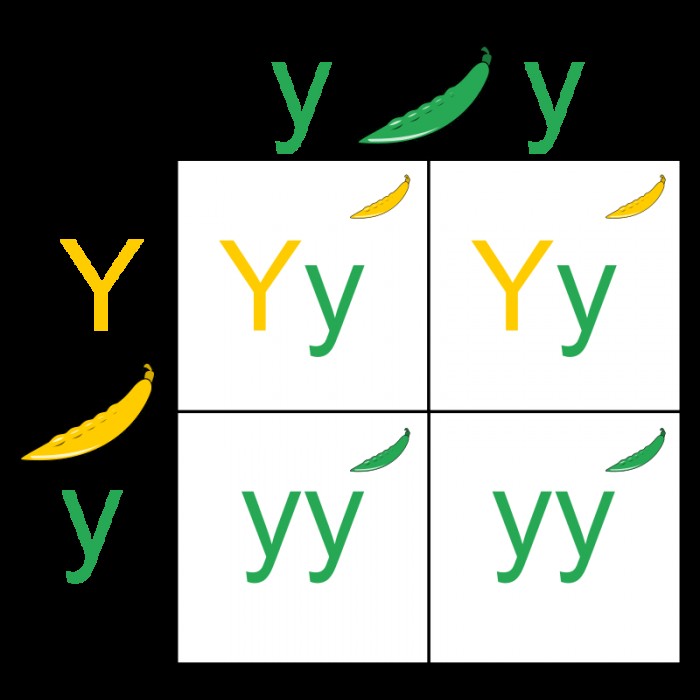
Punnett Square 이전에 상속 모델을 본 적이있을 것입니다. 이것은 유전자 상속이 두 가지 다른 대립 유전자라는 단순한 모델로서 지배적이고 열성입니다. 이 단순한 관계에서, 지배적 인 대립 유전자는 항상 유기체의 외관 또는 표현형에서 표현 될 열성 대립 유전자를 무시합니다. 지배적이고 열성적 인 특성에 대한이 이론은 그레고르 멘델 (Gregor Mendel)에 의해 만들어졌으며 부모의 특성이 단순히 자손 내에서 영구적으로 혼합 된 대중의 아이디어와 모순 되었기 때문에 중요했습니다. 그러나 이것은 당시 멘델이 믿었던 것에 대한 좋은 예 였지만, 현대 생물 학자들은 상속 이이 모델이 제안한 것만 큼 간단하지 않다는 것을 발견했습니다.
대립 유전자 쌍은 실제로 다양한 다른 지배 관계를 가질 수 있으며, 쌍 중 하나의 대립 유전자가 다른 대립 유전자가 적어도 완전히 보이지 않거나 표현되는 것을 막을 수 없을 수도 있습니다. 또한 특정 집단 내에서 주어진 유전자에 대해 때때로 많은 다른 대립 유전자가 있습니다.
Mendel은 식물과 완두콩 식물로 일련의 실험을 수행했습니다. 멘델은 자신의 관리에있는 완두콩 식물에 자주색 또는 흰 꽃이 있다는 것을 알았을 때 왜 이것이 왜 그런지 궁금해하기 시작했으며 왜 그들 중 누구도 혼합 된 라벤더 색상을 가지고 있지 않은 이유를 궁금해하기 시작했습니다. 멘델은 실험을 통해 두 가지 유형의 대립 유전자가 있다고 믿게되었습니다.
1900 년대 초, Carl Correns라는 이름의 독일 식물학자는 4시에 초점을 맞추었지만 멘델과 유사한 실험을 수행했습니다. Correns는 Mendel의 작업을 점프 오프 포인트로 사용했으며, 일반적으로 불완전한 지배력의 발견으로 인정받는 Correns였습니다. Correns는 자신의 치료에서 식물과 함께 일하는 동안 일부는 꽃잎에 혼합 된 색상을 표시한다는 것을 알았습니다. Correns는 모든 유전자형이 자체 표현형을 가질 가능성이 높았으며, 이들 중 다수에 대해 1 :2 :1 유전자형 비율이있을 가능성이 있다고 추론했다. 이 실현은 이종 접합 유기체가 단일 지배적 인 대립 유전자보다는 두 대립 유전자를 표시 할 수 있음을 의미했습니다.
유기체 대립 유전자 세트는 유전자형이라고합니다. 대립 유전자들 사이의 관계가 단순히 지배적이거나 자연적으로 열성이 아니라는 사실이 불완전한 지배적 인 상황에서 대립 유전자가 유기체의 표현형을 결정하지 않는다는 것을 의미하지는 않습니다. 대립 유전자는 여전히 표현형에 영향을 미치지 만,이 경우 다양한 대립 유전자는 특정 표현형을 생성하기 위해 고유 한 방식으로 서로 상호 작용할 수 있습니다. 경우에 따라, 이형 접합 유기체의 표현형은 부모의 표현형의 혼합 버전 일 수있다.
불완전한 지배력과 코다민
표현형의 블렌딩의 훌륭한 예는 antirrhinum majus라는 Snapdragon의 종입니다. 동형 접합 흰색 꽃과 동형 접합성 붉은 꽃이 DNA를 결합하면 분홍색 꽃을 생산합니다. 이것은 불완전한 지배의 예입니다. 안달루시아 닭에서 불완전한 지배력의 또 다른 예는 스페인 출신이며 깃털의 채색에 불완전한 지배력을 나타냅니다. 흑인 암컷 안달루시아 닭고기와 흰 수컷 닭 품종이라면 푸른 끈으로 깃털이있는 자손을 자주 생산합니다. 이것은 혼합 대립 유전자가 색소 멜라닌을 희석하고 깃털이 채색이 가벼워지는 방법을 반영합니다.
불완전한 지배력을 넘어서, 공동 지배라는 표현형 현상이 발생할 수 있습니다. 이 경우, 두 대립 유전자는 동시에 이형 접합 유기체를 Win으로 표현합니다. 실제로 MN으로 알려진 혈액형을 가진 사람들의 그룹이 있으며,이 혈액형을 결정하는 것은 특정 유전자의 대립 유전자입니다. l^m 대립 유전자가있는 사람은 적혈구 표면에 m 마커를 보여주고, l^n 마커를 가진 사람들은 다른 n 적혈구 마커를 보여줍니다. 동형 접합체 사람들은 적혈구에 두 마커 중 하나만 가지고 있지만, 이형 접합체는 그 중 두 마커를 나타냅니다. 두 표현형이 표시되는 공동 지배의 완벽한 예입니다.
많은 대립 유전자
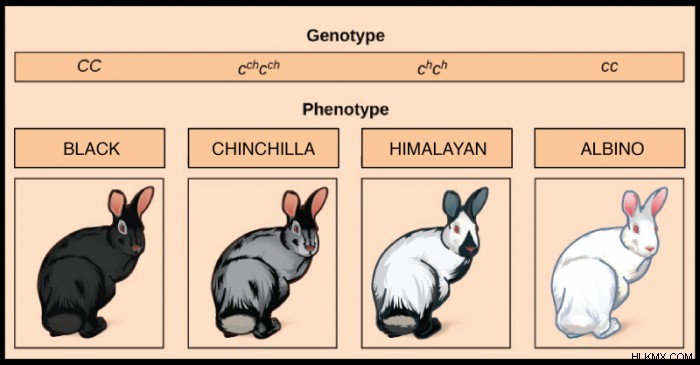
멘델의 연구는 모든 유전자에 대해 두 개의 대립 유전자 만 존재한다고 제안했을 수도 있지만, 실제로는 사실이 거의 없습니다. 이배체 유기체가 존재하지만, 다수의 대립 유전자는 종종 인구 수준에 존재하며 상기 집단 내의 다른 개체는 다른 대립 유전자 쌍을 가질 수있다. 예를 들어, 토끼는 종종 코트의 색을 정의하는 유전자의 4 가지 일반적인 대립 유전자를 갖습니다. 토끼에는 CC 쌍 (검은 색 또는 갈색 모피를 줄 수있는) 또는 CC 쌍 (흰 모피를 줄 수 있음)을 가질 수있을뿐만 아니라 C1C1 또는 C2 C2 쌍을 가질 수도 있습니다.
다처제 특성은 피부색, 체중, 높이 및 눈 색상과 같은 특성입니다. 또한, 다처제 특성은 다수의 유전자와 서로 다른 여러 대립 유전자의 상호 작용에 의해 결정된다. 이들 특성을 정의하는 역할을하는 유전자는 표현형에 대해 동등한 영향을 미치며 다른 염색체는 이들 유전자에 대한 대립 유전자를 함유한다. 다처제 특성에 영향을 미치는 대립 유전자의 효과는 본질적으로 부가 적이며, 이는 각각 다양한 양의 표현형 발현에 기여한다는 것을 의미합니다. 이 때문에 개인이 다양한 정도의 열성 표현형, 중간 표현형 및 지배적 표현형을 가질 수 있습니다.
이와 같은 다형성 상황에서 표현형이 표현되는 방식에 대해 생각할 수 있습니다.
더 지배적 인 대립 유전자를 물려받은 개인은 그 지배적 인 표현형의 더 큰 발현을 보여준다. 보다 열성적 인 대립 유전자를 물려받은 개인은 자연적으로 더 많은 열성 표현형을 나타냅니다. 마지막으로, 열성 및 지배적 대립 유전자의 조합을 물려받은 사람들은 다양한 지배적 및 열성 대립 유전자의 기여를 반영하는 표현형을 나타냅니다.