소개
키 큰 줄기 로셋 (일명 거대한 로제트)은 남미 북부 안데스, 동 아프리카 고원, 하와이와 카나리아 제도의 대형 화산을 포함한 많은 열대 산에서 높은 고도에서 자랍니다. 나무와 달리,이 식물의 우디 트렁크는 연간 성장 고리를 생성하지 않으므로 나이를 쉽게 결정할 수 없습니다. 대신, 줄기 높이의 변화는 일상적으로 플랜트 연령을 추정하기위한 대리자로 사용되었습니다. 신장률은 지난 50 년 동안 몇몇 줄기 로제트 종에 대해 측정되었다; 소규모 인구 또는 줄기로 세트 그룹에 대해 결정된 평균 성장률은 식물 연령을 계산하는 데 사용되었습니다. 이 식물들이 자라는 산 지역에 대한 접근이 어려워지고 다양한 물류 이유는 대부분의 성장 연구를 단기간으로 제한했으며, 종종 1 명 미만에서 몇 년 동안
안데스의 줄기로 세트는 남아메리카에서 접합자 (단수 :Frailejón,“Big Monk”)로 공동으로 공동으로 공장과 수도사에 대한 식물의 유사성으로 인해 공동으로 알려져있다. 연약함은 중앙 에콰도르에서 북부 베네수엘라에 이르기까지 ≥1200km 거리에있는 Andean Cordillera를 따라 발견됩니다 (Cuatrecasas, 2013). 여기에서보고 된 연구는 8 ° 52 'N에서 베네수엘라의 시에라 라 컬라 타 (Sierra La Culata)의 파라 모 피에드라 블랑 카스 (Páramo Piedras Blancas)의 키가 크고 풍부한 약한 연약한 종에 초점을 맞추 었습니다. 76 coespletia timotensis 에 대해 신장률이 측정되었습니다. 및 38 espeletia schultzii 10 년에서 ~ 20 년 사이의 기간 동안 4270-4525 미터 (~ 14010-14850 피트) 고도; 반복 사진의 도움으로 식물을 이전 하였다. 이 산맥에서 c. Timotensis 근처의 시에라 네바다 데 메다 (Sierra Nevada de Mérida)에서는 4780m (~ 15685 ')에서 발견되지만 최대 고도에 도달하지만, 근처의 시에라 네바다 데 메다 (~ 15685'); 실제로,이 식물은 안데스의 144 개의 연약한 종에 대해보고 된 가장 높은 고도에서 자랍니다 (Dizgranados, 2012). e. Schultzii , 연구 된 다른 식물 도이 범위에서 ~ 4300 미터 (~ 14110 ')까지 일반적입니다.
기후 맥락과 줄기 로스셋의 성장
상위 고도의 가혹한 기후 조건은 연약하게 자라는 연약한 모든 식물의 발달을 심각하게 제한합니다. 얼어 붙은 온도는 거의 매일 밤 발생하여지면 서리가 많이 발생하여 묘목의 대부분을 죽입니다. 평균 토양 온도가 0 ° C 아래로 떨어지는 낮은 영구 동토층 한계는 ~ 4550-4600m의 Piedras Blancas에서 발생합니다 (Schubert, 1975). 극한의 토양 가뭄은 또한 4 개월간의 건기 동안 발전하여 식물을 더욱 강조합니다. 북부 안데스에서 식물의 상한 경계는 열 결함과 생리 학적 건조로 인한 차갑고 건조한 한계입니다. 다수가 어려운 조건에도 불구하고 일부 연약한 크기로 자랍니다. c. Timotensis 종종 ~ 3.5m 높이 (사진)를 얻을 수 있습니다. e. Schultzii 훨씬 짧고 ~ 1.5m의 키에 도달합니다.


이 사진들은 c의 그룹을 보여줍니다. Timotensis ~ 4385 m에서; 오른쪽에있는 것은 Piedras Blancas에서 내가 찾은 가장 높은 로제트 중 하나입니다. 1982 년 1 월 10 일, 최상단 잎 팁까지의 높이는 291 ± 2 cm (사진 A)였다. 2000 년 2 월 29 일, 18 세, ~ 2 모 기간이 지난 후, 21cm에서 312 ± 2cm (사진 B)를 성장시켰다. ~ 1.16 cm/yr의 특정 성장률을 사용하면 (아래 토론 참조),이 식물에 대해 ~ 269 세의 추정됩니다. 물론 이것은 아마도이 식물의 생애 동안 균일 한 성장률을 잘못 가정 할 것입니다. 왼쪽에서 두 번째로 높은 장미는 같은 시간 간격 동안 훨씬 더 성장한 것 같습니다 (~ 28cm, ~ 1.54cm/yr). 스케일은 180cm 길이입니다.
짧은 시간에 걸친 초기 조사는 천천히 성장하는 개인들 사이에서 확장 속도의 상당한 변동성을 감지 할 수 없었지만, 1 ~ 20 년 동안의 신장을 평가하면 작은 변화가 극대화 될 수 있었고 식물 사이의 성장 차이는 ~ 34cm만큼 높았습니다. c에 대한 이전의 평균 성장. Timotensis 집단은 1.5 cm/yr, e에 대해 1.0 cm/yr로 추정되었다. Schultzii (Smith, 1981). 이 장기 연구 동안의 평균 트렁크 신장률은 크게 다르지 않았다 :각각 1.24 ± 0.46 cm/yr 및 1.04 ± 0.40 cm/yr. 그러나 놀랍고 전적으로 예상치 못한 결과는 이러한 평균 속도가 어느 종의 개별 식물들 사이에 광범위한 변화를 숨겼다는 것이었다.
.식물 성장의 변화의 원인
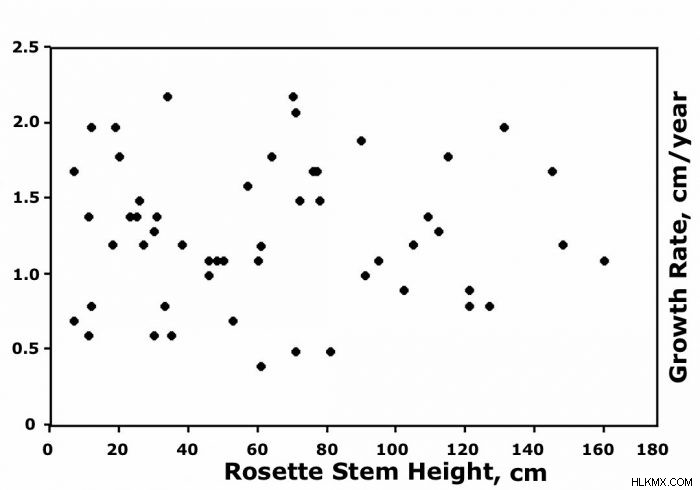
서로 가까이 자라는 식물조차도 비슷한 지형 위치와 토양 유형을 차지하고 동일한 측면으로 극심한 변동성을 나타 냈습니다. 그래프는 60 세의 생존에 대한 10 년, ~ 2 개월 동안 줄기 성장률 (cm/yr)이 인접한 c를 나타냅니다. Timotensis ~ 4285m의 계곡 바닥을 따라 ~ 0.4에서 2.5 cm/yr 사이의 변동이 발생했습니다. 남은 31 개의 남은 e에 대한 비슷한 산란. Schultzii 성장률은 ~ 0.4 ~ 2.4 cm/yr로 다양했습니다. 이 그래프는 또한 초기 식물 크기가 평균 성장과 관련이 없으며, 이는 임의적이고 식물 연령과 관련이없는 것처럼 보였음을 나타냅니다.
아마도, 개별 식물들 사이의 변화는 짧은 거리에 따라 변할 수있는 물리적 마이크로 사이트 특징의 차이 또는 특정 식물 근처의 생물학적 조건에서 발생할 수있다. 연약한 성장에 영향을 미치는 것으로 알려진 마이크로 사이트 요인은 토양 수분 함량, 토양 영양소 수준, 근처의 암석의 존재 및 인접한 식물 간의 경쟁적 상호 작용이 포함됩니다.
식물은 두 가지 방법으로 개별 줄기 로스셋의 성장에 영향을 줄 수 있습니다. 간호사 쿠션 식물에 대한 발아 또는 암석 옆에 연약한 발달이 촉진 될 수 있습니다. 둘 다 서리 주파수와 교란을 줄이고 일반적으로 토양 수분 조건을 개선합니다. 다른 상호 작용은 경쟁을 통해 성장을 억제 할 수 있습니다. 예를 들어, 사막 원주민 선인장에 대한 연구는 식물 바로 근처의 식생 풍부가 성장률에 부정적인 영향을 미치는“간섭 요인”을 생성한다는 것을 발견했습니다. 이 연구에서는 경쟁이 분석되지 않았지만, Páramo 식물에 대한 이전 데이터는 두 종의 식물 간섭 가능성을 강력하게 시사합니다. Timotensis 스탠드는 64%의 식물 덮개와 37 개의 혈관 식물 종이 풍부한 식생을 가졌다. e. Schultzii 스탠드는 또한 45 종의 식물과 61.5% 식물 덮개로 밀도가 높은 식물성이었다.
사망률에 의한 식물 감소
연구 기간이 끝날 때까지 충분한 식물이 생존하여 중요한 통계적 분석을 수행했지만 식물 사망률은 두 연약한 인구를 ~ 18-19%감소시켰다. c. Timotensis 묘목은 아마도 Frost Heaving 및/또는 반 차별 소에 의한 탐색에 의해 살해되었을 것입니다. 키가 크고 성숙한 c. Timotensis 이 종의 일반적인 사건 인 식물 토플 링에 의해 소멸되었다. 이것은 아마도 서리 교란으로 인한 경사면 이동 및 지상 이동과 관련이있을 것입니다. 토양 불안정성은 또한 키가 큰 Senecio keniodendron에 영향을 미칩니다 케냐 산 (아프리카)의 줄기 로셋은 가파르고 교란 된 경사에서 기울어 진“술 취한 스탠드”가 발생합니다 (Mahaney, 1980). 상당한 잎 사춘기와 수질 화합물의 높은 함량으로 인해 소는 먹는 것을 피하십시오. Schultzii ; 또한, 아마도 그들의 키가 짧기 때문에 아마도이 연약함은 기울어지는 것에 의해 실질적으로 영향을받지 않았으며, 대부분의 죽은 식물은 톱풀없이 서있었습니다.
.결론
결론적으로, STEM-ROSETTE 성장률, 인접한 식물의 심지어 심지어 평균 을 사용하여 6-16의 계수로 다양합니다. 날짜까지 개인 식물은 의미가없는 것 같습니다. 특정 식물에 대한 특정 성장 데이터가 부족한이 접근법은 미래의 연구에서 현명하지 않으며 중단되어야합니다. 어쨌든, Páramo Frailejones의 정확한 데이트에 대한 강조는 단지 부차적 일 수 있습니다. 이 장기 식물의 가장 주목할만한 측면은 그들이 적대적인 높은 안데스 환경에 얼마나 놀랍도록 적응했는지, 그리고 광범위한 산악 서식지를 점령하기 위해 어떻게 진화했는지가 될 수 있습니다. c. Timotensis , 다른 Andean Frailejón보다 더 큰 고도에서 성장하면, 높은 안데스 식물의 가장 두드러진 진화론 정점을 차지할 수 있습니다 (Cuatrecasas, 2013)
참조 :
- Cuatrecasas, J., 2013. 하위 트라이브 Espeletiinae (Heliantheae, Asteraceae)에 대한 체계적인 연구.
메모 뉴욕 식물원, 107, 1-689. - Dizgranados, M., 2012. 연약한 명칭 (Espeletiinae Cuatrec., Asteraceae).
phytokeys, 16, 1-52. https://phytokeys.pensoft.net/articles.php?id=1436. www.phytokeys.com. - Mahaney, W.C., 1980. 후기 4 차 암석 빙하, 케냐 산, 케냐 산. 저널 빙하,
25, 492-497. - Schubert, C. 1975. 베네수엘라 북서부의 빙하 및 주변 지형학
안데스. Eiszealter und Gegenwart, 26, 196-211. - Smith, A.P., 1981. 베네수엘라 안데스의 Espeletia (Compositae)의 성장과 인구 역학. Botany 48, 1-45에 대한 Smithsonian 기여.